



INTRODUCTION
The now-extinct aurochs (Bos primigenius), which ranged throughout much of Eurasia and Northern Africa, is widely accepted as the wild ancestor of modern domesticated cattle (MacHugh et al., 1998). Archaeological evidence shows that domestication of this animal occurred independently in the near East and in the Indian subcontinent between 10,000 to 8,000 yrs ago, giving rise to the two taxa of domestic cattle, namely B. taurus and B. indicus (Helmer et al., 2005; Bradley and Magee, 2006).
Genetic diversity of animal genetic resources (AnGR) compensates for changing environmental conditions, including climate, markets, and disease etc. Because AnGR are likely to continue to change and adapt in the future (Bruford et al., 2003; Toro and Caballero, 2005), the maintaining of their genetic diversity is important and essential for future use. However, during the last century, many indigenous domestic breeds became extinct by replacement or crossbreeding with exotic productive breeds. Therefore, the Food and Agriculture Organization of the United Nations (FAO) has encouraged a series of conservation measures designed to help prevent irreversible loss of domestic animal species (FAO, 2007b). In addition, conservation is one of the four Strategic Priority Areas of the recently adopted Global Plan of Action for AnGR (FAO, 2007a).
Molecular genetic studies within and across breeds are essential for the effective management of AnGR (Hall and Bradley, 1995; Ruane, 2000; Simianer, 2005). Mammalian mitochondrial DNA (mtDNA) shows several special features such as an absence of intron, maternal inheritance, the existence of single copy orthologous genes, lack of recombination events and a high mutation rate (Irwin et al., 1991; Pesole et al., 1999). Since the complete sequence of bovine mtDNA was published (Anderson et al., 1982), mtDNA have been widely used for genetic diversity and phylogenetic analysis among different cattle breeds (Bradley et al., 1996; Troy et al., 2001; Kikkawa et al., 2003; Kim et al., 2003; Berthouly et al., 2010; Mart├Łn-Burriel et al., 2011). Especially mtDNA cytochrome b (cyt b) has been broadly determined for the description of phylogenetic states of several species (Lau et al., 1998; Sultana et al., 2003; Souza et al., 2009; Stock et al., 2009; Yap et al., 2010).
More than 3,000 breeds of cattle are widely distributed throughout the world based on a Domestic Animal Diversity Information System (DAD-IS) from the FAO (http://dad.fao.org/). Four indigenous Korean cattle breeds are listed in the DAD-IS, and of them, the Chikso breed is an endangered breed with a unique brindle coat color. Efforts towards their conservation and proliferation have been made by government, universities and farmers. However, the genetic diversity, origin and evolution of this cattle breed have not been elucidated. The purposes of this investigation are to determine sequences of the mtDNA cyt b gene in full length, and to verify the genetic diversity, maternal origins and phylogenetic relationships of the Chikso breed.
MATERIALS AND METHODS
Sample collection and DNA extraction
The national management for the Chikso breed is carried out by 7 local institutions for AnGR management. The 239 blood samples were obtained from 5 institutions (Table 1) and were stored at ŌłÆ70┬░C until further processing. Genomic DNA was extracted from blood following an established protocol (Miller et al., 1988). The concentration of extracted DNA was measured using NanoDrop ND1000 (Thermo Scientific, USA), and was diluted to use as template for amplification.
Amplification and sequencing
The complete cyt b gene was amplified by using forward primer Bcyt-F: 5ŌĆÖ-TTCTTACATGGAATCT AACCATGA-3ŌĆÖ and reverse primer Bcyt-R: 5ŌĆÖ-GGGAGGTTAGTTGTTCTCCTTCTC-3ŌĆÖ. The forward and reverse primers were designed from tRNA-Glu and tRNA-Thr sequences of the mtDNA genome (GenBank accession no. V00654). Polymerase chain reaction (PCR) was carried out in a total volume of 25 ╬╝l, containing 10 ng of genomic DNA, 2.5 ╬╝l of 10├Śbuffer, 0.2 mM of dNTP, 10 pM of each primer and 1.5 units of Taq polymerase (TaKaRa, Japan). Thermal cycling was performed on a PTC-200 thermocycler (MJ Research Inc.) under the following conditions; 2 min denaturation at 94┬░C, followed by 35 cycles of 30 s at 94┬░C, 30 s at 60┬░C, 60 s at 72┬░C, and a final 5 min at 72┬░C before cooling to 4┬░C for 10 min. The amplified products were separated by electrophoresis on 1.2% agarose gels, and were visualized under UV illumination after staining with ethidium bromide. The PCR products were purified using a QIAquick PCR purification Kit (Qiagen, USA), and were directly sequenced on an ABI 3130xl Genetic Analyzer (PE Applied Biosystems, USA). The boundaries of the cyt b gene were determined by comparison with the B. taurus mtDNA genome sequence (GenBank accession no. V00654) as reported by Anderson et al. (1982).
Statistical and phylogenetic analyses
The sequences of the cyt b gene from different breeds were aligned in CLUSTAL W (Thompson et al., 1994). Numbers of nucleotide polymorphic sites (S) and haplotype (h), nucleotide diversity (Pi), haplotype diversity (Hd) and nucleotide divergence (Dxy) were performed in DNA sequence polymorphism Version 5.1 (Librado and Rozas, 2009). The Neighbor-joining (NJ) tree (Saitou and Nei, 1987) among haplotypes based on the cyt b gene sequences was reconstructed in MEGA 5.05 package (Tamura et al., 2011), with the reliability of the tree topology assessed by 1,000 bootstrap replications (Felsenstein, 1985). The NJ tree among breeds was constructed in MEGA 5.05 package on the basis of Dxy distances.
RESULTS AND DISCUSSION
Sequence composition and variation of the cyt b gene
The full-length coding sequences of the cyt b genes in 239 Chikso individuals were determined. All these sequences spanned 1,140 bp, started with an ATG translational start codon and ended with an AGA stop codon. No insertion/deletion or length variation was detected in these sequences. Compositional frequency of G base was the lowest (13.4%) other 3 bases (A, 31.2%; C, 30.2%; T, 25.2%). These patterns were very similar to those of a previous report which analyzed Chinese cattle breeds (Cai et al., 2007).
Fifteen polymorphic sites were identified among 239 cyt b coding sequences (Figure 1). All polymorphic sites were transition mutations, but there was no transversion mutation. Among the 15 polymorphic sites, 2 of them (site positions: 329, 643) were variant-singleton variable sites, and the others were two variant parsimony informative sites. In addition, 5 non-synonymous mutations (A232G, T329C, G643A, G736A, and A1114G) were identified, and substitutions of amino acid by these mutations were estimated as I78V, L110P, V215M, A246T, and I372V, respectively.
Thirteen haplotypes were defined by polymorphisms at 15 sites, and the cyt b sequences of these haplotypes were submitted to GenBank (Accession no. JX472262-74). The nucleotide diversity (Pi) of all individuals was 0.00055 and ranged from 0.00023 (GAN) to 0.00067 (CHU) (Table 1). Haplotype diversity (Hd) of Chikso was 0.4709 and ranged from 0.2577 (GAN) to 0.6114 (CHU). The Pi and Hd of the Chikso breed is lower than those of Chinese breeds of taurine lineage (Cai et al., 2007).
Haplotype distribution
For verification of the haplotype distribution of Chikso among Asian and European breeds, we obtained the cyt b gene sequences of Korean brown (called Hanwoo), Japanese black (JB), 6 Chinese breeds, and 3 European cattle breeds from GenBank database (Table 2). The distribution of the haplotypes in the Chikso and other breeds is shown in Table 3. Thirty five haplotypes (C1ŌĆōC35) were classified from 319 cyt b gene sequences. C2 was a major haplotype that contained 206 sequences (64.6%) from all breeds used. C1 was the second largest haplotype that included 54 sequences (16.9%). This haplotype did not contain any of the European breeds, nor did it contain 2 of the Chinese breeds (Kazakh and Jinnan). C3ŌĆōC13 haplotypes were Chikso-specific haplotypes, and there were no other breeds that contained at these haplotypes. C1 and C2 haplotypes contained 80.5% of cyt b sequences of Hanwoo, Yanbian, Zaosheng and JB breeds. Especially, all sequences of the Yanbian breed were contained at C1 and C2 haplotypes.
Phylogenetic analysis of the cyt b gene
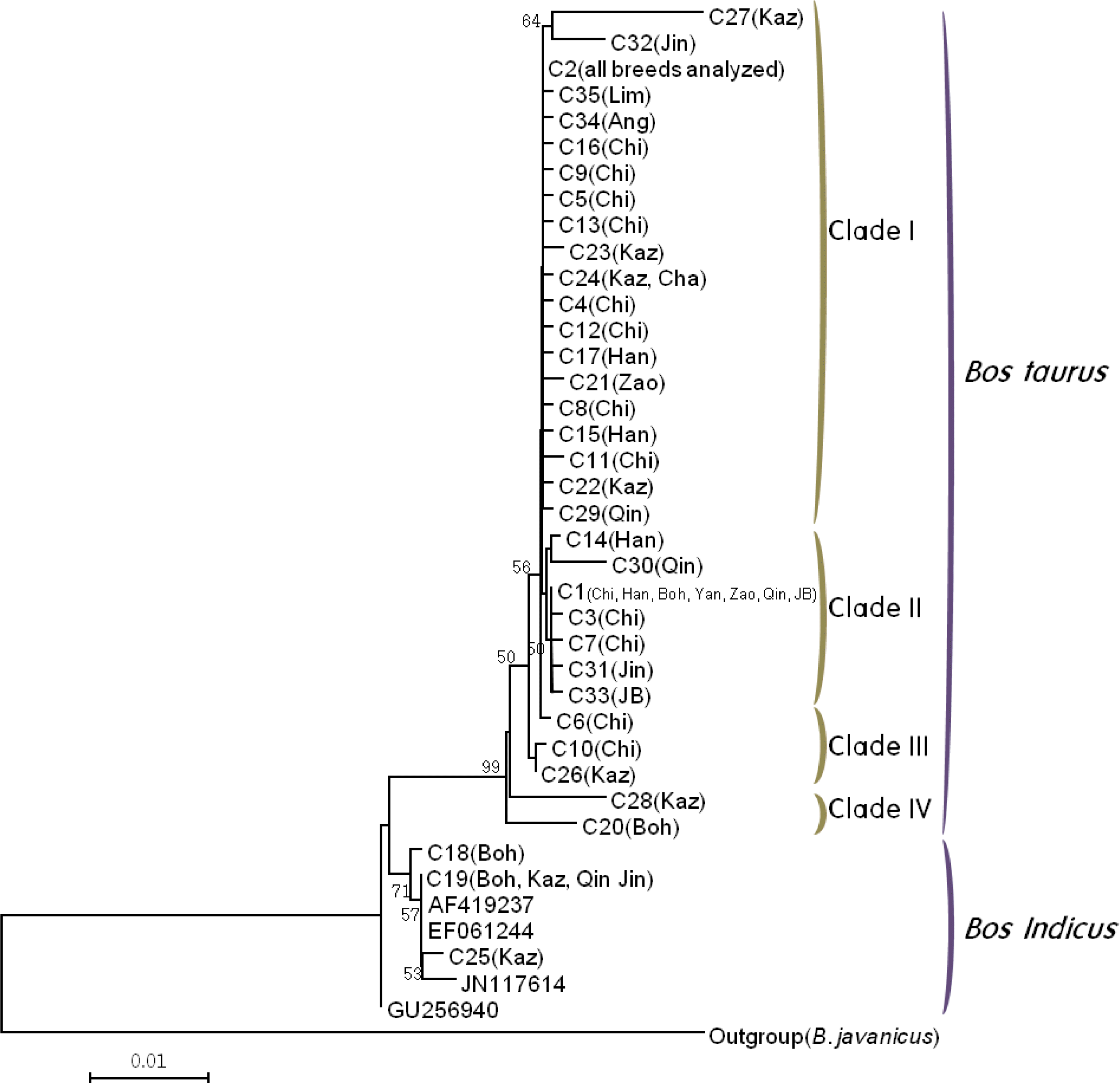
The neighbor-joining (NJ) tree was constructed to verify the phylogenetic relationship of 35 haplotypes of the cyt b gene, on the basis of Kimura two-parameter distances (Figure 2). Four cyt b sequences (GenBank Accession no. AF419237, EF061244, GU256940 and JN117614) from Zebu (B. indicus) were used together and B. javanicus was used as the outgroup (GenBank Accession no. D82889). The reliability of the tree topology was assessed by 1,000 bootstrap replications. The tree was divided into two distinct genetic lineages, B. taurus and B. indicus. Three (C18, C19 and C25) of 35 haplotypes belonged to B. indicus lineage and these haplotypes contained four Chinese breeds (Bohai black, Kazakh, Qinchuan and Jinnan). Cai et al. (2007) reported that 14 Chinese cattle breeds, including these four breeds, were of a mixture of two cattle lineages, B. taurus and B. indicus. Clade I consisted of haplotypes that derived from all breeds. Clades IIŌĆōIV were found only in Asian breeds and contained C1 haplotype. Clades III and IV contained Chikso and Chinese breeds. The Chikso breed was classified as being of B. taurus lineage and extended to Clade IŌĆōIII. The existence of a B. indicus lineage for Korean cattle breeds has not been found (Kikkawa et al., 2003; Kim et al., 2003; Mannen et al., 2004), and all Chikso individuals used in this study were verified as being of B. taurus lineage.
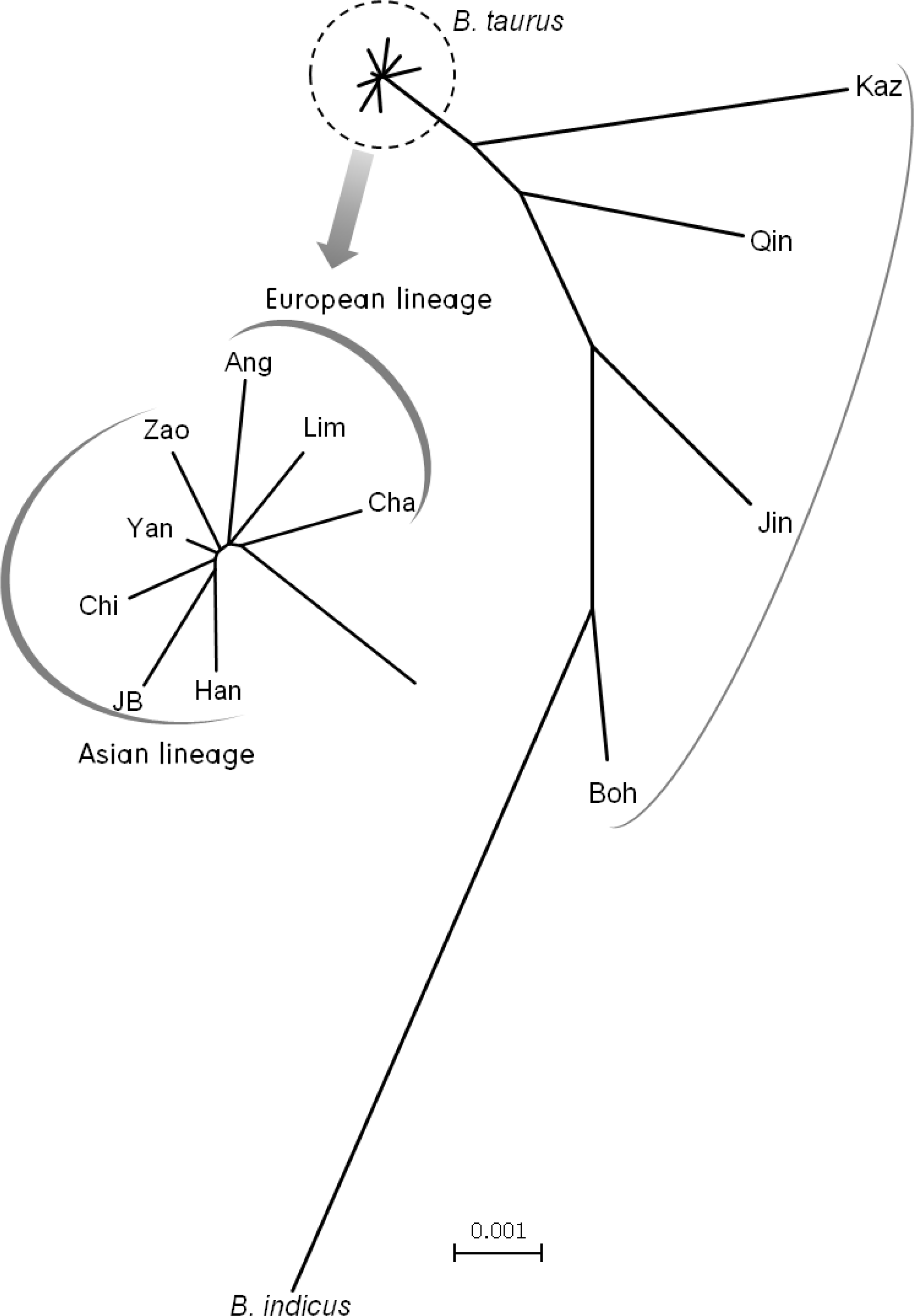
For verification of the phylogenetic status of the Chikso breed, a NJ tree was reconstructed based on Dxy genetic distances among Asian and European breeds (Figure 3). Four Chinese breeds, Bohai black, Jinnan, Qinchuan, Kazahk, were located in between B. taurus and B. indicus lineage. In the NJ tree based on 35 haplotypes (Figure 2), these breeds were verified to have two mtDNA maternal lineages, B. taurus and B. indicus. The phylogenetic location of these breeds was estimated to be due to these characteristics. Asian breeds and European breeds formed two independent groups in B. taurus lineage. Korean breeds, Chikso and Korean brown and Japanese black were located to the end of the tree through Chinese and European breeds from B. indicus. In the genetic distances estimated for verification of the genetic relationship between Chikso and other breeds, Chikso was more closely related to Yanbian (0.00040) than to Zaosheng (0.00064), Korean brown (0.00066) or JB (0.00066) (Table 4). These two Chinese breeds are distributed in north China (Cai et al., 2007). Yanbian breed samples were collected from Yanji City, which is the closest to the Korean peninsula. These results suggest that Korean cattle breeds might have been introduced from north China. This notion agrees with a report by Jia et al. (2010) that analyzed haplogroup patterns using mtDNA D-loop sequence.
Up until now, Chikso and Korean brown were classified as individual breeds on the sole basis of their coat color, however, there was no molecular genetic evidence. In this study, the Chikso breed was determined to be genetically closer to Yanbian and Zaosheng breeds than to the Korean brown breed in terms of phylogenetic analysis, although genetic distances showed slightly differing results. This result suggests that Chikso and Korean brown have a genetic difference based on their mtDNA cyt b gene, as well as based on their coat color for classification as breed.
In this study, we determined the full-length coding sequences of mtDNA cyt b gene and analyzed the genetic relationship and phylogenetic status of the Chikso breed. The Chikso breed was classified into B. taurus lineage and was found to be genetically more closely related to two Chinese breeds than to the Korean brown. For more detailed phylogenetic analysis and genetic characterization of the Chikso breed, additional genetic markers (e.g., mtDNA control region, Y chromosome and microsatellites) are necessary, as well as sample collection through the whole of Korea.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







