





INTRODUCTION
Melatonin (N-acetyl-5-methoxytryptamine) is an important hormone that is synthesized mainly in the pineal gland, and has a profound effect on serval physiology process including circadian rhythm and reproduction through its special receptors in birds [1ŌĆō3]. Three melatonin receptor subtypes, MTNR1A (alias MT1, Mel1a), MTNR1B (alias MT2, Mel1b), and MTNR1C (alias Mel1c), which belong to the superfamily of G protein-coupled receptors [4], have been cloned in birds [5,6].
In chicken, melatonin receptor subtypes were identified in ovaries [7,8], suggesting that melatonin directly affects ovarian function through activating of multiple receptors. In geese, the expression levels of MTNR1A, MTNR1B, and MTNR1C initially increased and later decreased during follicular development cycle, indicating that melatonin receptors participated in activating small white follicles and small yellow follicles to develop into subsequent greater hierarchical follicles [9]. In addition, in situ hybridization of MTNR1C mRNA combined with immunocytochemistry for gonadotropin-inhibitory hormone (GnIH), a key neurohormone controlling avian reproduction by inhibiting gonadal development [10], revealed a clear cellular colocalization of MTNR1C mRNA and GnIH in paraventricular nuclei [11]. It seems that melatonin most likely acts directly on GnIH neurons through MTNR1C to induce GnIH expression and regulate avian reproduction.
Shaoxing duck is a Chinese dominant layer breed, characterized by small body size, early maturity and high productivity. Modern Shaoxing duck achieve 50% egg production by 140 days of age, and the mean number of eggs at 500 days old was more than 300 [12]. Shaoxing ducks have been considered as a good source of duck eggs in China, and improving its egg production performance is of primary interest to breeders and farmers. The identification of single nucleotide polymorphisms (SNP) in candidate genes associating with economically important traits has become a powerful tool for genetic improvement of animal selection and production.
Numerous studies have investigated the relationship between melatonin receptor subtypes and egg production traits in different species, and made melatonin receptor genes potential candidate genes for QTLs [13,14]. However, whether or not nucleotide polymorphisms of MTNR1A, MTNR1B, and MTNR1C are associated with the egg production traits and egg quality traits in ducks is still unknown. Therefore, this experiment aimed to detect the SNPs in MTNR1A, MTNR1B, and MTNR1C and explore their associations with egg production traits and egg quality traits. The expression levels of these genes in ovary of ducks at different age at first egg were also determined.
MATERIALS AND METHODS
Performance traits and tissue collection
A total of 785 female Shaoxing ducks were randomly selected and raised in separate cages under similar environmental conditions and diet. The egg production traits of all ducks were recorded throughout the egg production process in terms of age at first egg, egg weight at 34 weeks old and total number of eggs at 34 weeks old and 72 weeks old. Egg collection for egg quality measurements took place at 34 weeks old. The following traits were recorded: egg shape index, shell thickness, shell strength, albumen height, Haugh unit score and eggshell color.
The blood samples of all the 785 individual ducks were collected from the wing vein using vacuum tubes containing dipotassium ethylene diamine tetraacetate (EDTA-K2) as an anticoagulant for further SNP analysis. To investigate the expression of the multiple forms of melatonin receptor mRNA, 6 ducks representing two groups (3 of each) according to the age at first egg (at 128 days of age or after 150 days of age) were carefully selected at age of 150 days. Ovaries were sampled after slaughter. These experiments were conducted in accordance with the Law of the PeopleŌĆÖs Republic of China on Animal Protection.
SNP discovery and genotyping
Based on the complete DNA sequence of Anas platyrhynchos genomic DNA sequence (NCBI accession no. NW_004676748.1), four pairs of primers were designed to amplify the target regions for SNP genotyping using the Primer Premier 6.0 software. Primer sequences are listed in Table 1.
Polymerase chain reaction (PCR) was carried out in a total volume of 15 ╬╝L consisting of 1 ╬╝L genomic DNA, 0.15 ╬╝L (10 ╬╝M) of each primer, 1.5 ╬╝L 10├Ś buffer, 1.5 ╬╝L (25 mmol/L) MgCl2, 0.3 ╬╝L (10 mmol/L) dNTPs, and 1.5 U Taq DNA polymerase. PCR conditions were as follow: 95┬░C for 3 min, followed by 35 cycles of 95┬░C for 15 s, 55┬░C for 15 s, 72┬░C for 30 s, and a final extension of 72┬░C for 10 min. The PCR products were sequenced commercially using ABI 3730XL automated sequencer (Applied Biosystems, Carlsbad, CA, USA) after being purified and extended. SNPs were identified by looking for multiple peaks at the same base pair.
Statistical analysis
The genotypic frequencies were calculated for each individual and the Hardy-Weinberg equilibrium was analyzed using the Chi-square test of PopGene Version 1.32. Pairwise tests for linkage disequilibrium (LD) were performed for each SNP using the SHEsisPlus online software platform (http://shesisplus.bio-x.cn/SHEsis.html). The egg production traits were compared among the genotypes. The association between the SNPs and different traits in 785 ducks were analyzed using SPSS 22 with the model Y = ╬╝+G+L+G├ŚL+e, where Y is the dependent variable (analyzed traits), ╬╝ is the overall mean, G is the genotype with a variation for the candidate gene, L is the fixed effect of breed, G├ŚL is the interaction between the genotype and duck population (a fixed effect), and e is the random error. The significance of the least squares means was tested with lest significance difference test.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from ovary samples using Trizol reagent according to the manufacturerŌĆÖs protocol. cDNA was synthesized from the total RNA according to the manufacturerŌĆÖs protocol of TransScript First-Strand cDNA Synthesis SuperMix (TransGen, Beijing, China). Quantitative real-time PCR was performed using ABI 7500 (Applied Biosystems, Foster City, CA, USA). The mRNA expression levels were determined by Applied Biosystems real-time PCR using the MTNR1A, MTNR1B, and MTNR1C mRNA-specific primers and the ╬▓-acting gene as the internal control (Table 1). The 20 ╬╝L amplification reaction contained 10 ╬╝L of SYBR Green Universal PCR Master Mix, 2 ╬╝L cDNA, 0.4 ╬╝M of each primer and nuclease free water up to 20 ╬╝L. Thermal parameter used to amplify the template started with an initial denaturation at 94┬░C for 3 min followed by 40 cycles of 94┬░C for 10 s and annealing at 60┬░C for 30 s. The relative expression levels of the genes test were calculated using the 2ŌłÆ╬öCt method. Groups with different letters were significantly different at p<0.05.
RESULTS
Genetic polymorphism of melatonin receptor genes
Seven SNPs were identified in all three melatonin receptor gene exons respectively by direct sequencing (Figure 1), their corresponding allele and genotype frequencies are presented in Table 2. Hardy-Weinberg equilibrium tests showed that the alleles of MTNR1A g. 268C>T, MTNR1C g. 10C>T, and MTNR1C g. 108C>T were in Hardy-Weinberg equilibrium (p>0.05), and others deviated from the Hardy-Weinberg equilibrium (p<0.05) (Table 1). The LD tests of Shaoxing duck population showed that two SNPs (g. 41C>T, g. 161T>C) identified in MTNR1B gene were completely linked (DŌĆ▓ = 1.0 and r2 = 1.0) (Supplementary Table S1), providing only three possible haplotypes.
Association of SNPs with egg production traits and egg quality traits
The association between the SNPs and egg production traits are shown in Table 3. For the MTNR1A g. 268C>T locus, age at first egg of the genotype CT was significantly less than that of the genotype TT (p<0.05). The g. 108C>T locus of MTNR1C was significantly associated with age at first egg and total number of eggs during 34-week egg-laying period (p<0.05). For MTNR1C g. 108C> T, the egg number of the genotype CT at 34 weeks old was significantly higher than that of the genotype CC (p<0.05), and ducks with the genotype TT exhibited significantly earlier age at first egg than that of the genotype CC (p<0.05). No significant association was found among other SNPs and egg weight, total number of eggs at 34 weeks old, total number of eggs at 72 weeks old or age at first egg (p>0.05).
As shown in Table 4, there were no significant association of 7 SNPs with egg quality trait. Furthermore, relation of SNPs with the color of eggshell was analyzed using chi-square test. When white eggshell was compared with blue eggshell, no significant differences in genotypes of MTNR1B or MTNR1C were observed except MTNR1A (p>0.05). The genotype TT and CT of g. 268C>T in MTNR1A were more common in Shaoxing ducks laying white eggshell compared with blue eggshell (Supplementary Table S2), suggesting MTNR1A g. 268C>T SNP may affect the eggshell color.
Expression of melatonin receptor mRNA in ovary
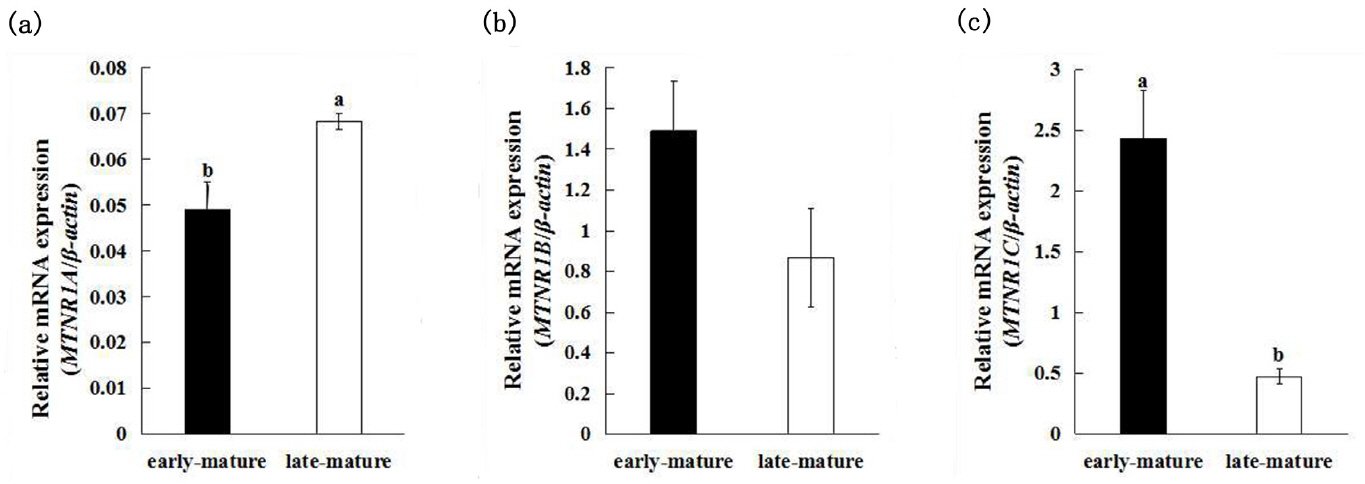
The expression of MTNR1A, MTNR1B, and MTNR1C between early-mature (first egg at 128 days of age) group and late-mature (first egg after 150 days of age) group showed different patterns (Figure 1). The late-mature group had significantly higher MTNR1A mRNA expression (0.0684┬▒0.0017) compared to the early-mature group (0.0491┬▒0.0060) (p<0.05) (Figure 2a), whereas there was no significant difference of MTNR1B mRNA expression between the early-mature group (1.491┬▒0.4580) and the late-mature group (0.869┬▒0.2435) (p> 0.05) (Figure 2b). The MTNR1C expression level of ovary was significantly higher in the early-mature group (2.4328┬▒0.3936) than in late-mature group (p<0.05) (0.4756┬▒0.0595) (Figure 2c).
DISCUSSION
Age at first egg, an important trait indicating sexual maturation and egg production performance, was negatively correlated with number of eggs [15ŌĆō17]. However, age at first egg was controlled by polygenes [17] with low to moderate estimated heritability ranging from 0.13 to 0.20 [18ŌĆō20], making the conventional breeding method ineffective. Therefore, molecular assisted selection becomes a powerful tool for improving egg production traits and raising economic benefits.
As previously reported in other avian species, melatonin regulates gonadal maturation by suppressing luteinizing hormone (LH) secretion, and stimulating GnIH and gonadotropin releasing hormone secretion [21ŌĆō24]. MTNR1C mRNA colocalized with GnIH neurous [11] indicating that melatonin participates in gonadal maturation through binding its receptors. Therefore, melatonin receptors would be possible markers for selecting an early maturing breed. Although there were many studies seeking correlations between markers of candidate genes and age at first egg, such as growth hormone, prolactin, neuropeptide Y, follicle-stimulating hormone receptor, and SH3-domain GRB2-like 2 [25ŌĆō29], little was known regarding polymorphisms in melatonin receptors of duck.
Our study demonstrated that g. 268C>T of MTNR1A gene shows a strong association with age of first egg. Beside, g. 268C>T of MTNR1C gene was found to be associated with total number of eggs at age of 34 weeks and age at first egg. A study in chickens also showed that SNPs (JQ249890:g. 384T>C, JQ249891:g. 387 T>C) locating at MTNR1A and SNP (JQ249896:g. 294 G>A) locating at MTNR1C were significantly linked with age at first egg [14]. The novel SNPs which was founded in our study, indicated that melatonin receptor genes may affect age at first egg and involve in sexual mature of ducks.
The exact role of melatonin receptors in different species during the reproductive process is not well understood. It was reported that melatonin receptor density showed a striking downregulation in brain when songbirds were less than fully reproductively mature, and subsequent resumed during photorefractory state [30]. While Abd et al [31] reported that MTNR1B and MTNR1C expression increased accompanying a delay in sexual maturity of Japanese quails. In our study, the expression of MTNR1C mRNA in late-mature group was significantly decreased compared to the early-mature group. Summarizing the results of the above researches, it is reasonable to suggest that MTNR1C activates avian sexual mature by down-regulating its expression level. The MTNR1A mRNA expression in our experiment showed a contrary tendency with MTNR1C mRNA, suggesting melatonin receptors regulate ovarian function through different mechanisms. Further studies are continuing to elucidate the exact role of melatonin receptors in avian sexual maturity.
Collectively, the results of present study suggest that the MTNR1A and MTNR1C genes play an important role in egg production of ducks, but also SNPs in these genes could be used as markers in molecular marker-assisted selection for duck reproduction traits.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print







