





INTRODUCTION
The intestinal microbiota is a complex ecosystem that plays a major role in the physiology and health of the host [1]. Studies have found that host genetics impact the composition of gut microbiota and their metabolites [2]. Pig is one of the most important economic animals and an ideal model for studying human physiological function and disease [3]. Pigs of different breeds have phenotypically different genetic makeups that result in diverse physiological traits as well as gastrointestinal microbiomes [4]. The Saba pig is a traditional native pig breed found in Chuxiong Autonomous Prefecture, Yunnan Province, and has a high reproductive rate, good meat quality, and the capacity for high utilization of coarse feed [5]. However, large-scale domestic commercial farms rarely feed the local breed because of their unfavorable characteristics, which include slow growth rate and low lean meat rate. Cross breeding has been widely used in pig breeding because of pig heterosis and breed complementarity [6].
Commercial pig producers generally use a terminal crossbreeding system with three breeds, and, at present, a three-way cross model is largely used in commercial pig production [7]. Among the pigs involved in this model, Duroc pigs and Berkshire pigs are world-renowned lean pig breeds and are the main terminal sire breeds in commercial pig production [8].
An investigation into the intestinal microbial composition of hybrid pigs would reveal and promote the utilization of the genetic characteristics of different pig breeds. Thus, in the present study, we performed 16S rRNA gene sequencing analysis of the gut microbiomes of Duroc×(Berkshire×Saba) (DBS) pig and Berkshire×(Duroc×Saba) (BDS) pigs. Our aim was to characterize the differences in gut microbiome composition between these two hybrids and evaluate the underlying association of the gut microbiota with growth performance.
MATERIALS AND METHODS
Animal ethics statement
All animal works performed were approved by the Institutional Animal Care and Use Committee of the Yunnan Agricultural University (No. YNAU20181006). The care and use of animals fully complied with local animal welfare laws, guidelines and policies.
Experimental animal and sample preparation
The experiment was carried out on a commercial farm in the city of Kunming, China. We selected 100 35-day (d)-old DBS and 100 BDS three-way hybrid weaned piglets (half males and half females), and all piglets were raised in the same temperature-controlled house, with 10 piglets per pen and 10 pens per treatment (length 4 m, width 3 m). There is a central passage with pens on either side, the pen floor was made of concrete and the house is naturally ventilated. Pigs were fed a three-stage NRC diet (Table 1), and were allowed free access to water and were ad libitum fed till 210 d.
At the end of the 210-day trial, 20 pigs (ten from each group) were sacrificed by exsanguination. Then the cecal chyme samples were collected into sterile tubes and quick-frozen in liquid nitrogen before stored at −80°C for 16S rRNA analysis. Body weight gain, feed consumption, and the feed/gain (F/G) ratio for these pigs were recorded from 35 d to 210 d. Polymerase chain reaction amplification, 16S rRNA amplicon sequencing, and processing of sequence data were performed as previously described [9].
Statistical analysis
Experimental data, including those for growth performance and microbial abundance, were analyzed using the SPSS 22.0 software (IBM SPSS Statistics for Windows; IBM Corp, NY, USA). The Shapiro-Wilk test was used to evaluate normality. The general linear model of the Duncan multiple comparison test was used to analyze the parameter data, and Kruskal-Wallis analysis of variance was used for the phylum and genus level microbiota. Growth performance and microbial data are expressed as the mean±standard error. Phylogenetic studies of the PICRUSt communities were based on operational taxonomic units (OTUs) [10]. To assess the correlation between dominant genera and growth performance, we performed Spearman’s test in GraphPad Prism 7.0 [11]. A p-value <0.05 was considered to be statistically significant.
RESULTS
Comparison of growth performances of DBS and BDS hybrid pigs
The final body weight (FBW) and average daily gain (ADG) of DBS pigs were significantly higher compared to those of BDS pigs, while the F/G ratio of DBS pigs was significantly lower than that of BDS pigs (p<0.05; Table 2).
Changes in the diversity of the microbial community in the cecum
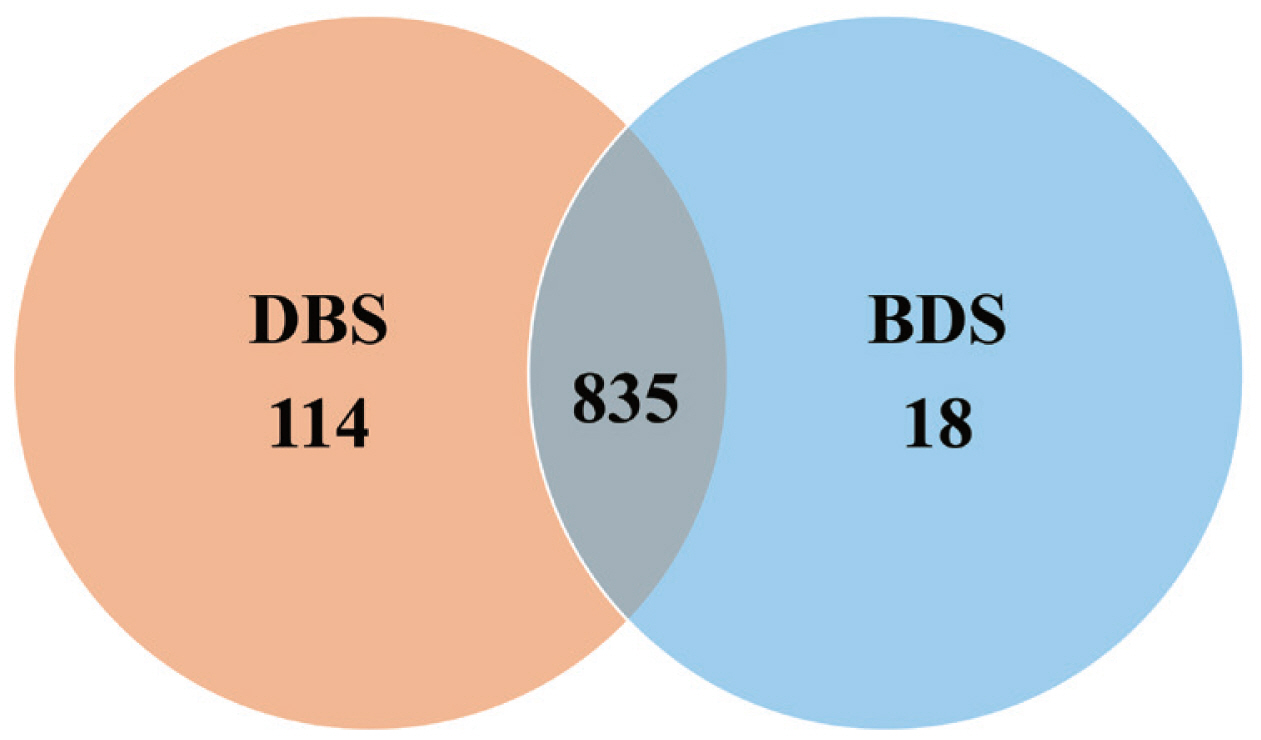
A total of 20 cecal samples generated 1,209,608 clean reads, with an average of 60,480 clean reads per sample and an average clean read length of 415 bp. OTUs were obtained at a sequence similarity level of 97%. A Venn diagram was used to reveal the shared and unique microbial percentages in the DBS and/or BDS groups. Additionally, 835 core OTUs were identified in the two groups, and 114 and 18 unique OTUs were identified in the DBS and BDS groups, respectively (Figure 1).
Diversity indices were calculated based on the OTUs of each library. The Chao1 index, phylogenetic diversity whole tree index, good’s coverage, Shannon index, observed species indices, and Simpson index were used to evaluate the abundance and diversity of the microbial species in the samples. We found that there were differences in the alpha diversity indices between DBS and BDS pigs, but the differences were not statistically significant (Table 3). Principal component analyses (PCA) were used to estimate the beta diversity between the two groups. The PCA plot of the unweighted unifrac distances showed that the DBS and BDS groups had a distinct difference (p<0.05; Figure 2).
Microbial composition in the ceca of DBS pigs and BDS pigs
At the phylum level, 16 and 14 taxa were identified in DBS and BDS pigs, respectively. The dominant phyla of the two groups were Bacteroidetes, Firmicutes, Spirochetes, and Proteobacteria, and the abundances of these 4 dominant phyla in DBS pigs were 55.23%, 36.65%, 2.7%, and 2.86%, respectively. Similarly, the abundances of these 4 dominant phyla in the BDS pigs were 59%, 34.86%, 2.92%, and 2.27%, respectively (Table 4).
Twenty-one genera were identified in the DBS and BDS groups. The abundances of Prevotella, Roseburia, and Anaerovibrio in DBS pigs were significantly lower than those in BDS pigs (p<0.01). The abundances of Bacteroides, Methanomassiliicoccus, and Parabacteroides in DBS pigs were significantly higher than those in BDS pigs (p<0.05). The abundances of Eubacterium and Clostridium XI in DBS pigs were significantly higher than those in BDS pigs (p<0.01; Table 5).
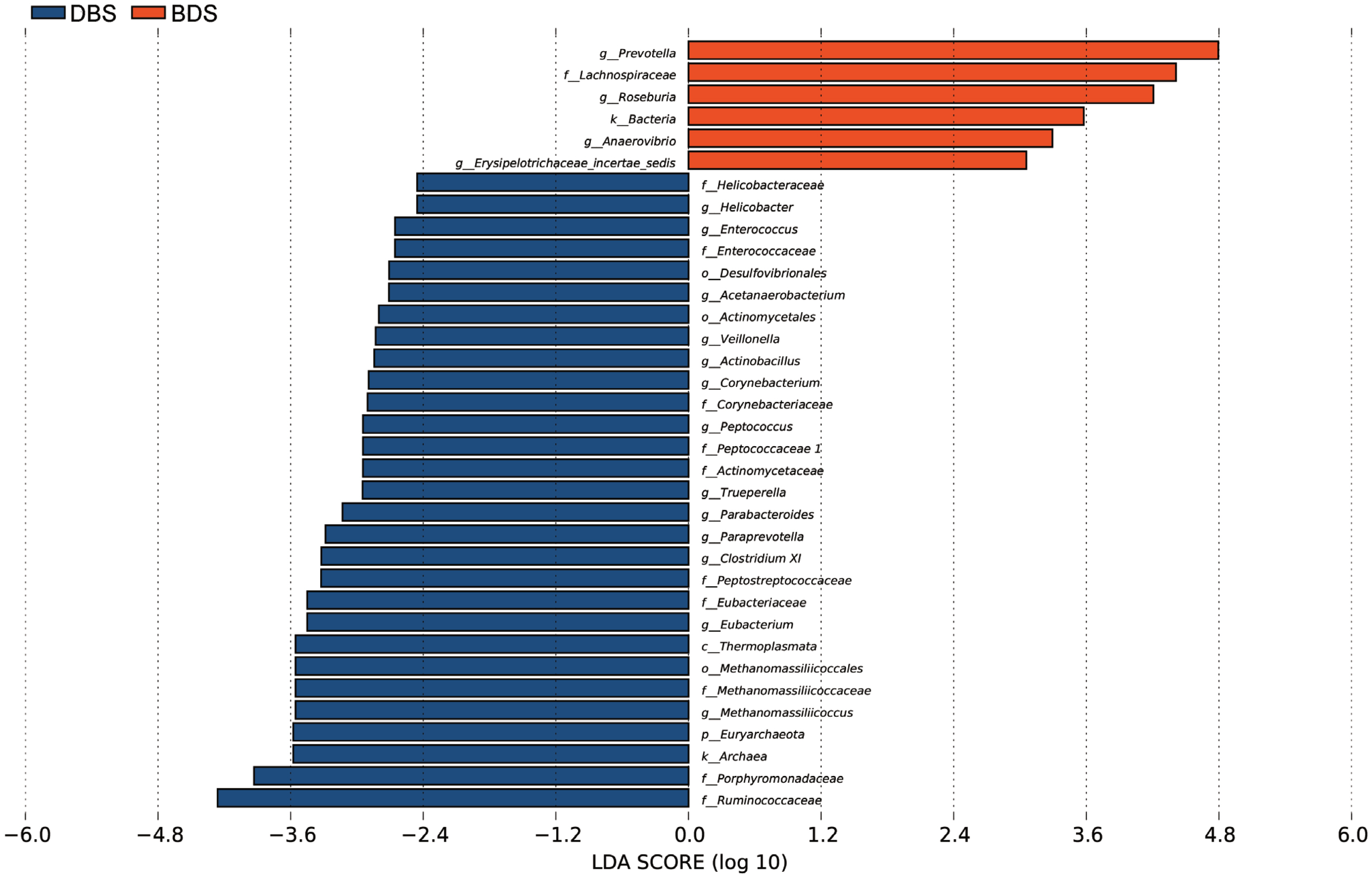
To determine the specific bacterial taxonomic groups associated with different hybrid pigs, we performed a linear discriminant analysis effect size (LEfSe) to compare the cecal microbiotas of DBS and BDS pigs. The data showed that at the kingdom level, Archaea was significantly enriched in the DBS group, and Bacteria were significantly enriched in the BDS group (Figure 3). At the phylum level, Desulfovibrionales was significantly enriched in the DBS group. At the class level, Thermoplasmata was significantly enriched in the DBS group. At the order level, Methanomassilicoccales, Desulfovibrionales, and Actinomycetales were significantly enriched in the DBS group. At the family level, there was significant enrichment of Helicobacteraceae, Enterococcaceae, Corynebacteriaceae, Methanomassiliicoccaceae, Porphyromonadaceae, Ruminococcaceae, Peptococcaceae I, Actinomycetaceae, Peptostreptococcaceae, and Eubacteriaceae in the DBS group and that of Lachnospiraceae in the BDS group. At the genus level, Helicobacter, Enterococcus, Acetanaerobacterium, Veillonella, Actinobacillus, Corynebacterium, Eubacterium, Methanomassiliicoccus, Peptococcus, Trueperella, Parabacteroides, Paraprevotella, and Clostridium XI were significantly enriched in the DBS group, and Prevotella, Roseburia, Anaerovibrio, and Erysipelotrichaceae incertae sedis were significantly enriched in the BDS group.
Correlation between microbiota and growth performance
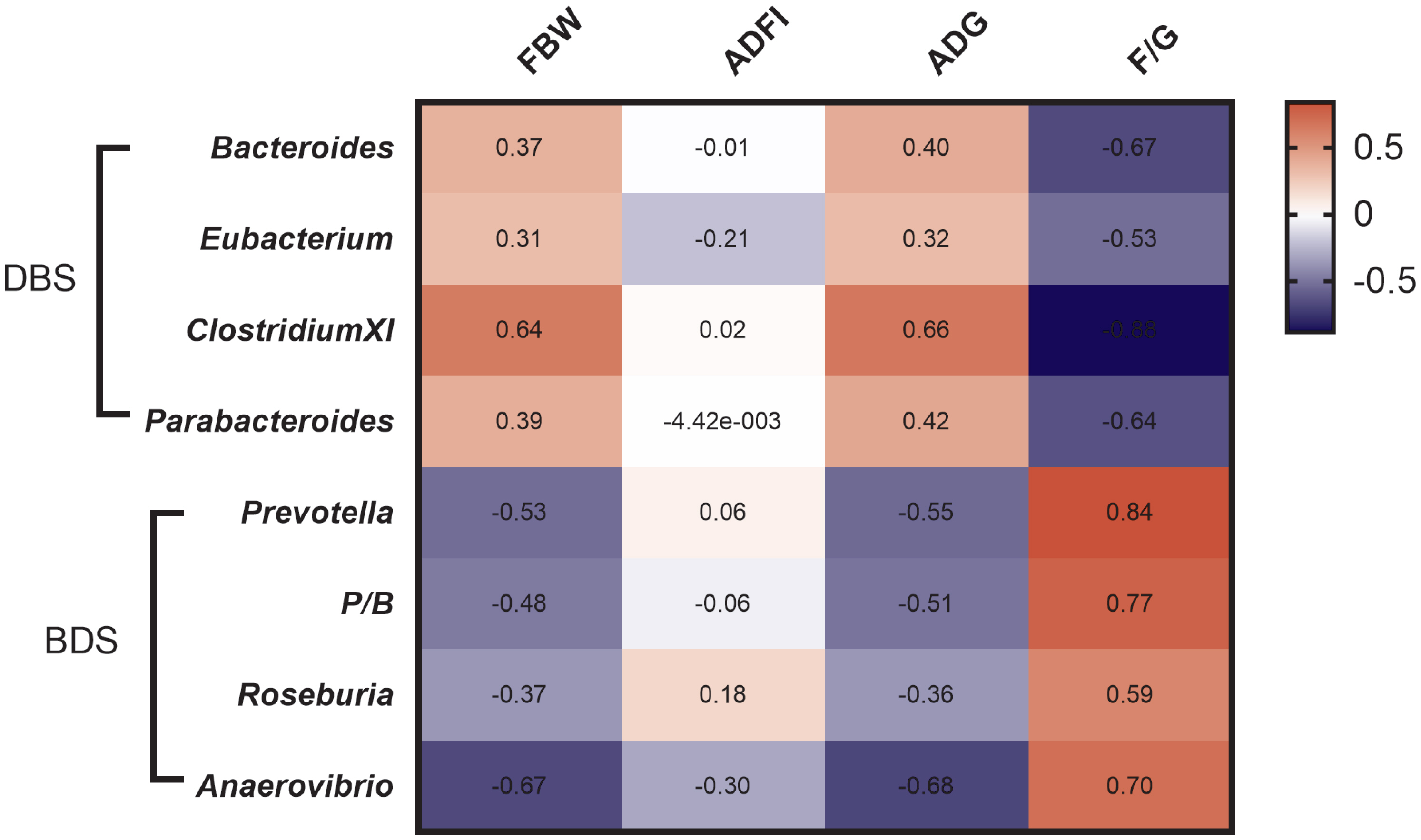
To investigate the correlation between cecal microbiota and the growth performance of these two hybrid pig models, a heat map was constructed (Figure 4). Our data showed that the FBW and ADG were positively correlated with Bacteroides, ClostridiumXI, and Parabacteroides but negatively correlated with Prevotella, Prevotella/Bacteroides (P/B) ratio, Roseburia, and Anaerovibrio. The F/G ratio was positively correlated with Bacteroides, Eubacterium, ClostridiumXI, and Parabacteroides but negatively correlated with Prevotella, P/B ratio, Roseburia, and Anaerovibrio.
DISCUSSION
In the modern pig industry, crossbreeding is an effective method to improve the efficiency of and profit from production [12]. The Berkshire breed has excellent meat quality features, such as thin muscle fiber and excellent water holding capacity [13], the Duroc breed has both excellent growth rate and intramuscular fat and is used as a terminal sire when fattening pigs are produced [14]. Previous studies have reported that the growth rate of pigs produced by crossing DDL (Duroc×(Duroc×Landrace) and BDL (Berkshire×(Duroc× Landrace) pigs was higher than those of pure-breed Berkshire and Duroc pigs [15]. Studies have also shown that the ADG of Hampshire and Landrace hybrid pigs increased significantly and that the growth performance of hybrid pigs was higher than that of purebred pigs [16]. Further, studies have found that hybrid pigs show superior genetics at the production level [17]. The Saba pig is one of the most important local pig breeds in Yunnan Province and the main maternal breed used in hybrid systems in central Yunnan. Our results show that the FBW and ADG of DBS pigs were significantly higher than those of BDS pigs, while the F/G ratio of DBS pigs was significantly lower than that of BDS pigs. These data show that DBS pigs have a better growth performance than BDS pigs.
Intestinal microbiota are generally thought to play a major role in bodily functions and have a considerable impact on the growth and health of the host [18,19]. Furthermore, studies have shown that host genetics have a significant impact on the composition of the intestinal microbiome [20]. Moreover, the type of pig breed is a major factor affecting the intestinal microbial composition. An investigation of the jejunal and colonic microbial communities in pure-breed Meishan and Yorkshire piglets showed that the type of breed has a significant impact on the bacterial community structure on days 14 and 49 [21]. In addition, the importance of host genetics in shaping the gastrointestinal microbiota in hybrids has also been demonstrated in mouse models. For instance, researchers found that hybrid mice displayed widespread transgressive phenotypes in their bacterial communities, which exhibited a structure associated with aberrant immune gene expression and increased intestinal pathology [22]. Therefore, host genetics also have a major impact on the microbiome and metabolites in hybrid animals [2]. Our data showed that the gut microbiota differed between two related hybrid pigs, with differences existing in some genera; no difference existed in the microbial diversity between DBS and BDS pigs, suggesting that hybridization has a major effect on the cecal microbiota composition at the genus level.
Many studies on intestinal microbiota have shown that Firmicutes and Bacteroides are the two most dominant phyla in the animal intestine [23], which is consistent with our work showing that the dominant intestinal microbes in the two hybrid pigs were Firmicutes and Bacteroides at the phylum level. Bacteroides and Prevotella are the two main genera in the Bacteroidetes phylum [24]. Prevotella ferments complex polysaccharides from the diet to produce succinate [25]. Bacteroides produce propionate and harvest energy more efficiently from food than commensal gut microbes [19,26]. Additionally, high fat and protein intake are associated with increased levels of Bacteroides, while high fiber intake is associated with increased levels of Prevotella [24]. A previous study showed that overweight pregnant women had more Bacteroides-Prevotella group bacteria than normal-weight pregnant women [27]. Individuals with a high P/B ratio lost more body weight and body fat than individuals with a low P/B ratio, confirming that individuals with a high P/B ratio are more susceptible to weight loss [28]. Our results showed that FBW and ADG are positively correlated with Bacteroides and negatively correlated with Prevotella and P/B ratio, which agrees with previous studies. These observations suggest that the P/B ratio may be considered as an important biomarker in weight management as well as pig breeding.
Taken together, the experimental findings presented in this study show that DBS pigs have better growth performance than BDS pigs. In addition, differences existed in some genera, with no difference in diversity between DBS and BDS pigs. Furthermore, our study provides evidence that that host genetics affect the cecal microbiota composition and the porcine gut microbiota is associated with growth performance, thereby suggesting that gut microbiota composition may be a useful biomarker in porcine genetics and breeding.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







