





INTRODUCTION
Adiponectin, an adipokine belonging to the complement factor C1Q protein family [1], is expressed in liver sinusoid endothelial cells, skeletal muscle and bone-forming cells as well as in adipocytes [2ŌĆō4]. Adiponectin, which is expressed by the AdipoQ gene, consists of four parts: an amino-terminal signal peptide, a variable region, a collagenous domain and a carboxyl-terminal globular domain [5]. There are two receptors for adiponectin, AdipoR1 and AdipoR2. Both are seven-transmembrane domain proteins with an intracellular N-terminus and highly expressed in skeletal muscle (AdipoR1) and adipose tissue (AdipoR2) [6]. Adiponectin circulates in the plasma in several forms: globular adiponectin (gAd) interacts with AdipoR1; whereas full-length adiponectin (fAd) binds to both AdipoR1 and AdipoR2 to regulate glucose and fatty acid metabolism [7].
Emerging evidence suggests that adiponectin enhances glucose uptake by increasing GLUT4 translocation to the cell membrane [8] and promotes fatty acid oxidation by activating adenosine 5ŌĆ▓-monophosphate (AMP)-activated protein kinase (AMPK), p38-mitogen-activated protein kinase, and peroxisome proliferator-activated receptor alpha pathways [9,10]. In addition, adiponectin inhibits gluconeogenesis by phosphorylating AMPK and acetyl-CoA carboxylase (ACC) [11]. For example, intravenous infusion of adiponectin in mice leads to lower expression of PEPCK and G6Pase in the liver, which inhibits endogenous glucose production [12]. In myocytes, fAd increases the number of mitochondria and enhances fatty acid oxidation by activating CaMKK╬▓ and PGC1-╬▒ [13]. Studies have also reported that overexpression of the AdipoQ gene in fat tissue and fibroblasts results in increased lipid accumulation [14]. Therefore, the results of these studies suggest that adiponectin plays important roles in regulating glucose uptake and fatty acid metabolism.
It has been shown that ACC and fatty-acid synthase (FASN) are two key enzymes in de novo fatty acid synthesis. ACC couples with FASN to increase fatty acid production in Escherichia coli and humans [15,16]. In addition, sterol regulatory element-binding protein-1 (SREBP-1), CCAAT/enhancer-binding protein-╬▒ (C/EBP╬▒) and peroxisome proliferator-activated receptor ╬│ (PPAR╬│) are master adipogenic transcription factors that have been proven to promote the formation of white adipose tissue [17]. As previously demonstrated, PPAR╬│ and SREBP-1 cooperatively play key roles in regulating the redifferentiation of bovine adipofibroblasts [18]. Moreover, C/EBP╬▒ has important functions in regulating cell proliferation and differentiation, especially adipocyte differentiation [19], and PPAR╬│ and C/EBP╬▒ together induce adipogenesis and adipocyte differentiation [20]. These studies demonstrate that these genes play pivotal roles in fatty acid synthesis and adipocyte differentiation. Although adiponectin has been shown to regulate fatty acid metabolism, it remains unknown whether adiponectin regulates the expression levels of these adipogenic-related genes.
An increasing number of studies on adiponectin are being carried out in humans and mice, but information on the gene encoding adiponectin in domestic animals is still limited. In addition, the role of adiponectin in the adipogenic differentiation of muscle satellite cells is unknown. In this study, we cloned and characterized the goat AdipoQ, AdipoR1, and AdipoR2 genes and detected their mRNA expression levels in different tissues. In addition, we quantified the expression levels of these adipogenic-related genes in skeletal muscle satellite cells (SMSCs) overexpressing adiponectin. The results of this study provide information on the expression patterns of the AdipoQ, AdipoR1, and AdipoR2 genes in goat as well as the functions of adiponectin in regulating fatty acid synthesis.
MATERIALS AND METHODS
Animals and tissue collection
All procedures were performed according to the guidelines established by the Sichuan Province Committee for the Protection of Laboratory Animals. Eleven tissues, namely, the ovary, spleen, rumen, liver, brain, heart, gastrocnemius muscle, biceps femoris muscle, longissimus dorsi muscle (LD), and mesenteric and subcutaneous adipose tissue, were dissected from three three-day-old female Nanjiang brown goats. All tissues were immediately frozen in liquid nitrogen and then stored at ŌłÆ80┬░C.
RNA isolation and cDNA synthesis
Total RNA was extracted by using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturerŌĆÖs protocol. The integrity of total RNA was examined by agarose gel electrophoresis, and the purity and concentration were determined using a NanoDrop (Thermo Fisher, Waltham, MA, USA) instrument. First-strand complementary DNA (cDNA) was synthesized using 2 ╬╝g of total RNA and a PrimeScript RT Reagent Kit (TaKaRa, Tokyo, Japan) according to the manufacturerŌĆÖs protocol.
Primer design and gene cloning
Based on the conserved regions of the human and cattle AdipoQ, AdipoR1, and AdipoR2 gene sequences, primer pairs for gene cloning and quantitative polymerase chain reaction (qPCR) were designed by Primer Premier 5.0 (Table 1) and synthesized by Shanghai Sangon Biotechnology Company (Shanghai, China). The total reaction volume was 10 ╬╝L and included 1 ╬╝L cDNA, 5 ╬╝L 2├Ś Taq PCR Master Mix, 0.8 ╬╝L (8 pM) primer pairs (ADF/R for AdipoQ, ADR1F/R for AdipoR1 and ADR2F/R for AdipoR2) and 3.2 ╬╝L ddH2O. The PCR procedures were as follows: initial denaturation at 95┬░C for 3 min; 35 cycles of 95┬░C for 30 s, optimum annealing temperature (Table 1) for 30 s and 72┬░C for 1 to 2 min; and a final extension at 72┬░C for 5 min. All products were separated by agarose gel electrophoresis, purified using a gel extraction reagent kit (Tiangen, Beijing, China), cloned into the pMD19-T vector (TaKaRa, Japan) and sequenced by Shanghai Invitrogen Biotechnology Company (Shanghai, China).
Semi-quantitative reverse transcription-PCR analysis
To detect the mRNA tissue distributions of the AdipoQ, AdipoR1, and AdipoR2 genes, we performed semi-quantitative reverse transcription (RT)-PCR. The total reaction volume was 20 ╬╝L and included 2 ╬╝L cDNA, 10 ╬╝L 2├Ś Taq PCR Master Mix, 1.2 ╬╝L (12 pM) primer pairs (ADQF/R for AdipoQ, ADR1QF/R for AdipoR1 and ADR2QF/R for AdipoR2) and 6.8 ╬╝L ddH2O. The PCR procedure was as follows: initial denaturation at 95┬░C for 3 min; 26 to 32 cycles at 95┬░C for 30 s, optimum annealing temperature (Table 1) for 30 s and 72┬░C for 10 s; and a final extension at 72┬░C for 5 min. All products were examined by agarose gel electrophoresis to estimate mRNA abundance in the 11 tissues tested; ╬▓-actin was used as an internal control gene.
Quantitative real-time PCR
qPCR was carried out using a SYBR Green-based kit in 10-╬╝L reaction volumes containing 5 ╬╝L SYBR Green real-time PCR Master Mix (TaKaRa, Japan), 0.8 ╬╝L cDNA and 0.4 ╬╝L each primer; a Bio-Rad CFX96 qPCR instrument (Bio-Rad, Hercules, CA, USA) was used. The qPCR procedure was as follows: initial denaturation at 95┬░C for 3 min; 40 cycles at 95┬░C for 30 s, alternative annealing for 20 s, and 72┬░C for 15 s; and a final extension for 5 min. Melting curve analysis was used to confirm specific PCR products. Gene expression levels were quantified relative to ╬▓-actin expression using the comparative Ct (2ŌłÆ╬ö╬öCt) value method.
Cell culture
SMSCs were isolated using 1 mg/mL Pronase E (Sigma, St. Louis, MO, USA) from the longissimus dorsi muscles from 3-day-old female Nanjiang brown goats. After enzymatic digestion, cells were separated from tissue fragments by repeated centrifugation at 500├Śg for 5 min, followed by filtration through a 200-╬╝m filter and then a 50-╬╝m Nytex filter. SMSCs were cultured in DulbeccoŌĆÖs modified EagleŌĆÖs medium (DMEM) containing 10% (v/v) fetal bovine serum, 100 U/mL penicillin, and 0.1 mg/mL streptomycin under conditions of 5% CO2 and 37┬░C. When the cells had grown to 80% to 90% confluence, adipogenic differentiation of satellite cells was induced by transferring the cells into DMEM supplemented with an isobutylmethylxanthine, dexamethasone and insulin (MDI) cocktail (0.5 mM 1-methyl-3-isobutylxanthine, 1 ╬╝M dexamethasone, and 10 ╬╝g/mL insulin), followed by supplementation with 10 ╬╝g/mL insulin for 1 day. The medium was replaced every two days. Cells were visualized by staining with oil red O solution.
Plasmid construction and transient transfection
Using Primer Premier 5.0, a primer pair (Hind III-F, BamH I-R) was designed to amplify the coding region of AdipoQ for plasmid construction. PCR products and the pEGFP-N1 (Clontech, San Francisco, CA, USA) vector were digested with Hind III and BamH I restriction enzymes (TaKaRa, Japan), purified with a gel extraction reagent kit and ligated using T4 DNA ligase (TaKaRa, Japan). The resulting plasmid (pEGFP-N1-AD) was sequenced to confirm the correct orientation and reading frame.
Transient transfection was performed according to the manufacturerŌĆÖs protocol: 4 ╬╝g plasmid and 10 ╬╝L Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) were mixed, cultured for 20 min and then transfected into cells. Twenty-four hours after transfection, the cells were rinsed with phosphate-buffered saline; a portion was fixed with 4% formaldehyde for 30 min at room temperature to detect the cellular distribution of the adiponectin fusion protein using a fluorescence microscope (Nikon, Tokyo, Japan); other cells were suspended in TRIzol reagent for RNA extraction and expression analysis. The results were normalized to the expression of ╬▓-actin, and the expression of the control group (pEGFP-N1) was set as 1 for expression analysis of each gene. The relative mRNA expression analysis was performed using SAS 9.0 software with the generalized linear model process. DuncanŌĆÖs new multiple range test was applied to analyze statistical significance.
Immunocytochemistry
Expression of adiponectin in SMSCs was detected by immunocytochemical staining. Briefly, cells were fixed with 4% paraformaldehyde for 30 min and then permeabilized in 0.25% Triton X-100 with shaking at room temperature for 30 min, followed by blocking with 1% bovine serum albumin for 1 h. The cells were then incubated overnight at 4┬░C with an anti-adiponectin primary antibody (1:100) (Boster Company, Wuhan, China). The secondary antibody, horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G (1:200), was incubated with the cells for 30 min. Cells incubated without the primary antibody were used as the negative control.
Bioinformatic analysis
ORF Finder (http://www.ncbi.nlm.nihgov/gorf/gorf.html) and DNASTAR were used to predict the open reading frames of the three genes. The domains of the three proteins were predicted by ProtParam (http://cn.expasy.org/tools/protparam.html) and SMART (http://smart.embl-heidelberg.de/), and the functional domain diagram was generated with DOG 2.0 software [21]. Amino acid sequences were aligned using ClustalX (http://www.ebi.ac.uk/clustalw/).
RESULTS
Molecular characterization of AdipoQ, AdipoR1, and AdipoR2
The full-length coding sequences of the goat AdipoQ, AdipoR1, and AdipoR2 genes were obtained (accession numbers: JX573539, KC286912 and JX573540). The coding regions of the AdipoQ, AdipoR1, and AdipoR2 are 720 bp, 1,128 bp, and 1,176 bp, respectively. The AdipoQ gene encodes a 239-aa protein with a molecular weight of 25.95 kDa. The AdipoR1 gene encodes a 375-aa protein with a molecular weight of 42.41 kDa, and the AdipoR2 gene encodes a 391-aa protein with a molecular weight of 43.81 kDa. These three genes share high similarity with other mammalian orthologous genes (Supplementary Table S1). Goat AdipoR1 shows the highest homology with other mammals, with the amino acid sequence sharing more than 96% identity with other mammalian sequences. Goat AdipoQ and AdipoR2 exhibit relatively high homology with other mammalian sequences. The amino acid sequence of AdipoQ is 81.17%, 87.45%, 81.59%, and 99.16% identical to those of Homo sapiens, Bos taurus, Mus musculus, and Ovis aries, respectively. The amino acid sequence of AdipoR2 shows similar identity to those of humans (86.27%), Bos taurus (92.49%), Mus musculus (84.20%), and Ovis aries (94.04%). Prediction of the deduced protein structure showed that adiponectin harbors a 137-aa C1Q domain; although the receptors both contain a 220-aa seven-transmembrane domain, these receptors are not identical (Figure 1).
Tissue distribution of goat AdipoQ, AdipoR1, and AdipoR2
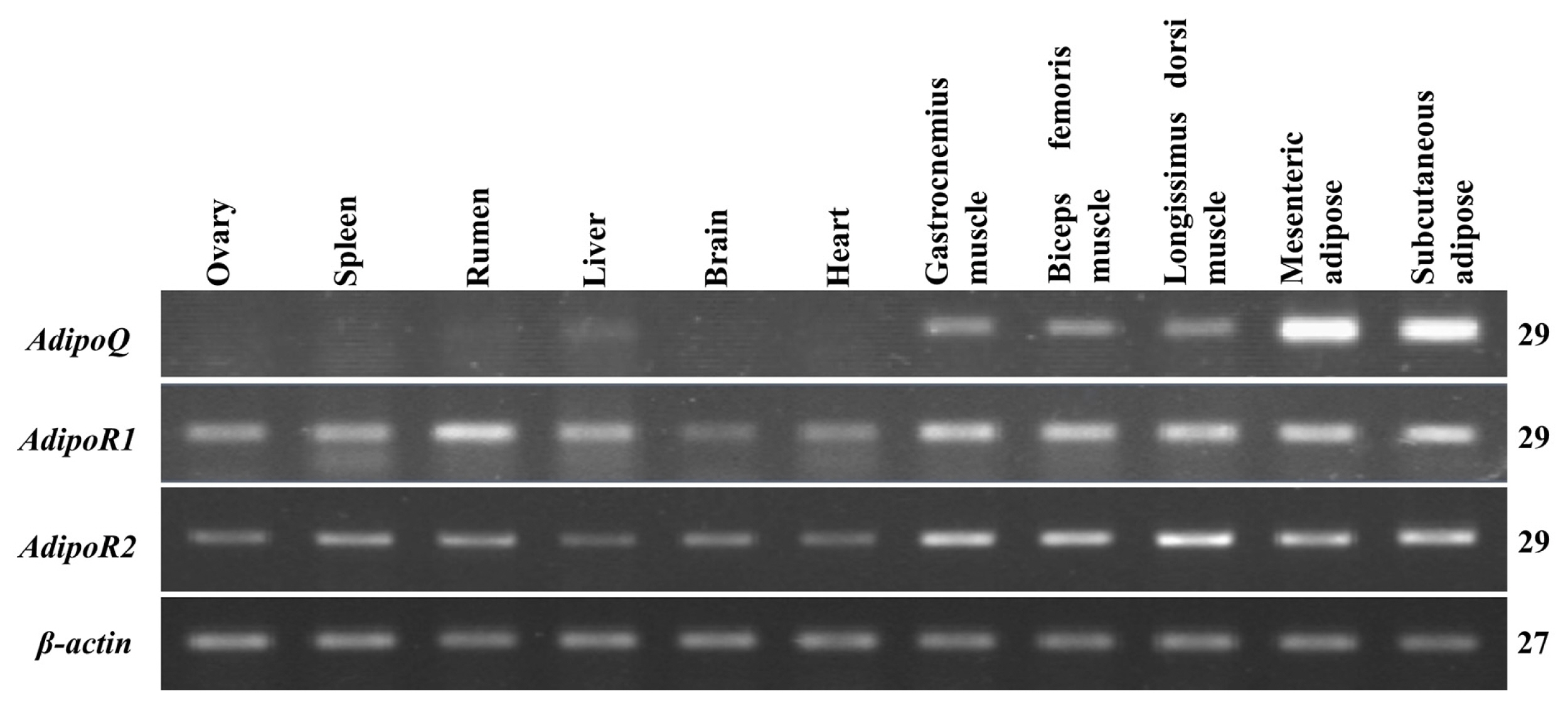
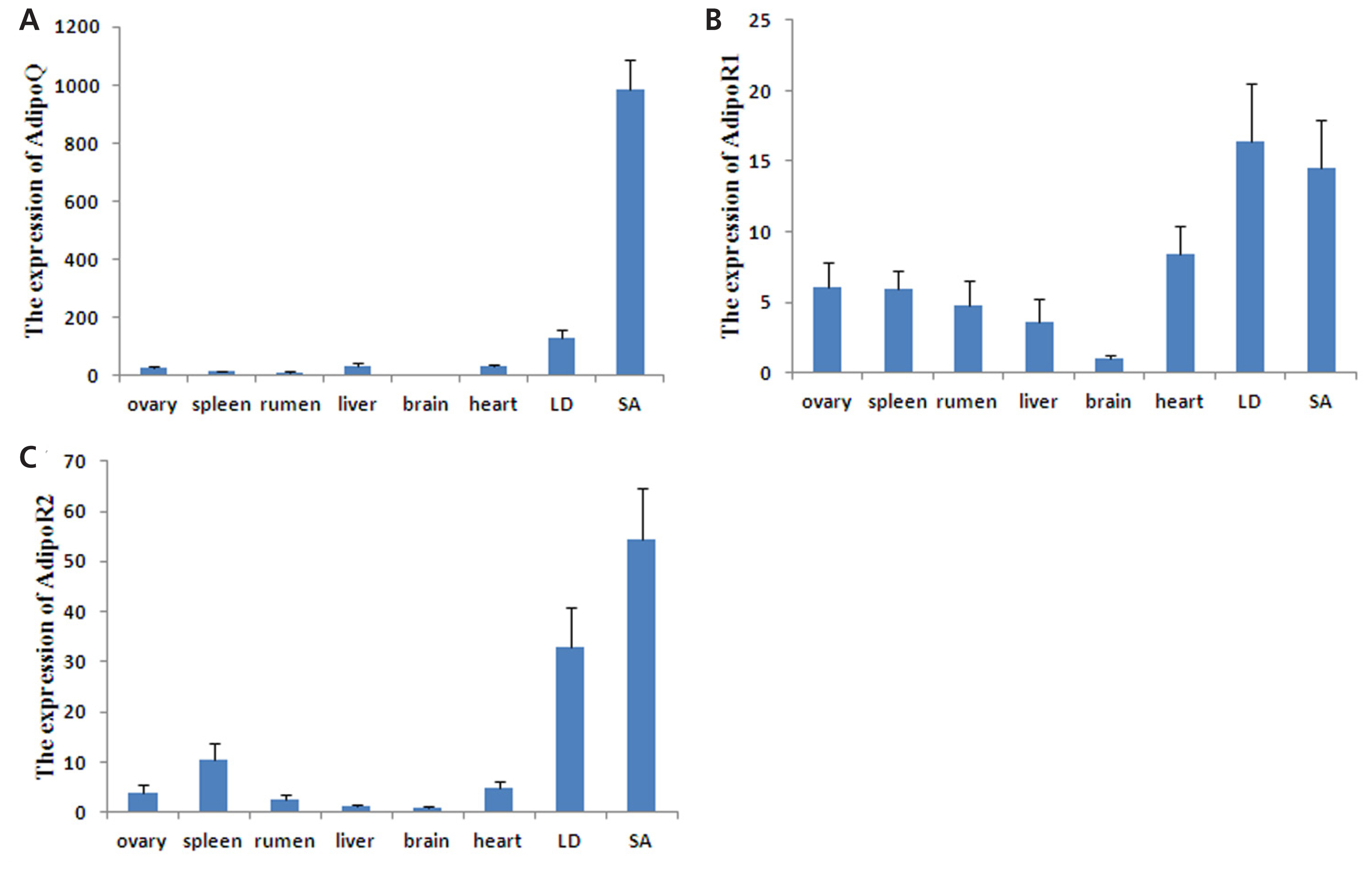
The mRNA tissue distribution of these three genes was detected by semi-quantitative RT-PCR, and the agarose gel electrophoresis results are shown in Figure 2. AdipoQ expression was predominantly observed in mesenteric and subcutaneous adipose tissues but was weakly expressed in the gastrocnemius muscle, LD muscle and biceps femoris muscle; expression was not detected in other tissues. AdipoR1 mRNA was found to be expressed in almost all tissues, with high expression in skeletal muscles and the rumen, followed by adipose tissue; its expression in other tissues was relatively weak. Similarly, AdipoR2 was expressed in all tissues but was most abundant in adipose and skeletal muscle tissues; it was weakly expressed in other tissues. The expression patterns of these three genes were further quantitated by qPCR (Figure 3). AdipoQ mRNA was predominantly expressed in adipose tissue, whereas its expression in skeletal muscles was relatively weak. AdipoR1 and AdipoR2 mRNA were expressed in all tissues examined, with the highest expression in adipose and skeletal muscle tissues.
Subcellular localization of goat adiponectin in SMSCs
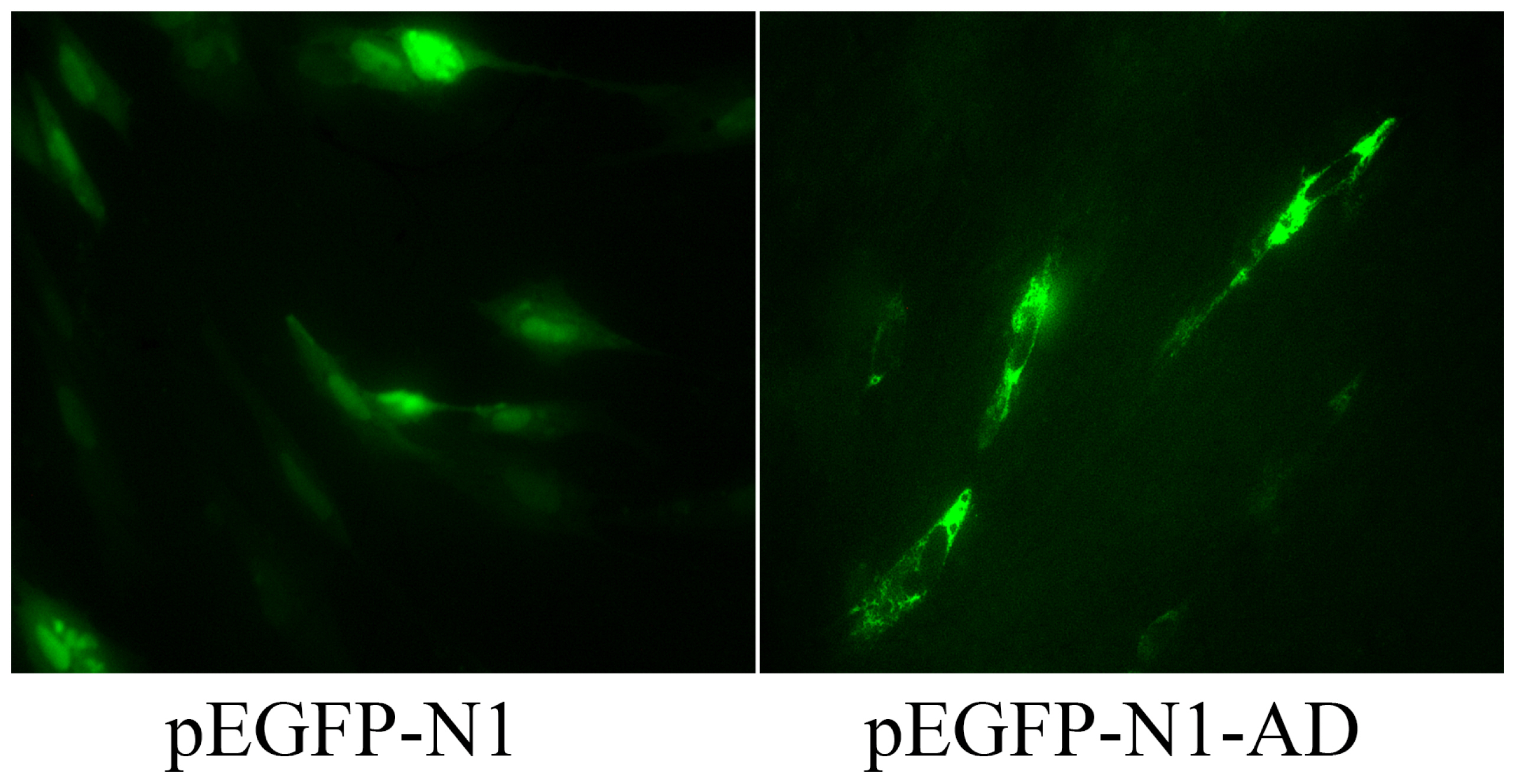
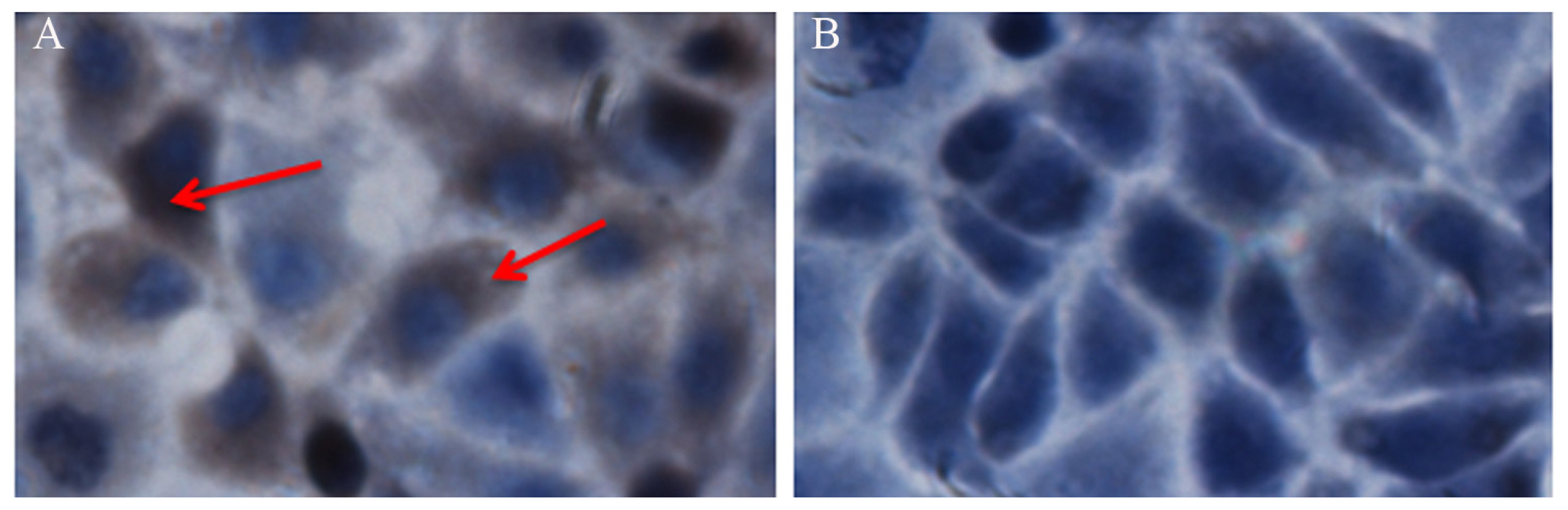
Fluorescence analysis showed a green fluorescent signal throughout cells of the control group transfected with the pEGFP-N1 plasmid; In contrast, the green fluorescence of the adiponectin fusion protein was only observed in the cytoplasm of cells transfected with the pEGFP-AD plasmid (Figure 4), suggesting that adiponectin is localized to the cytoplasm. Furthermore, immunostaining in SMSCs using an antibody against the human adiponectin protein revealed strong positive staining within the goat SMSC cytoplasm (Figure 5A) compared with the controls (Figure 5B).
Adiponectin promotes fatty acid synthesis in skeletal muscle satellite cells
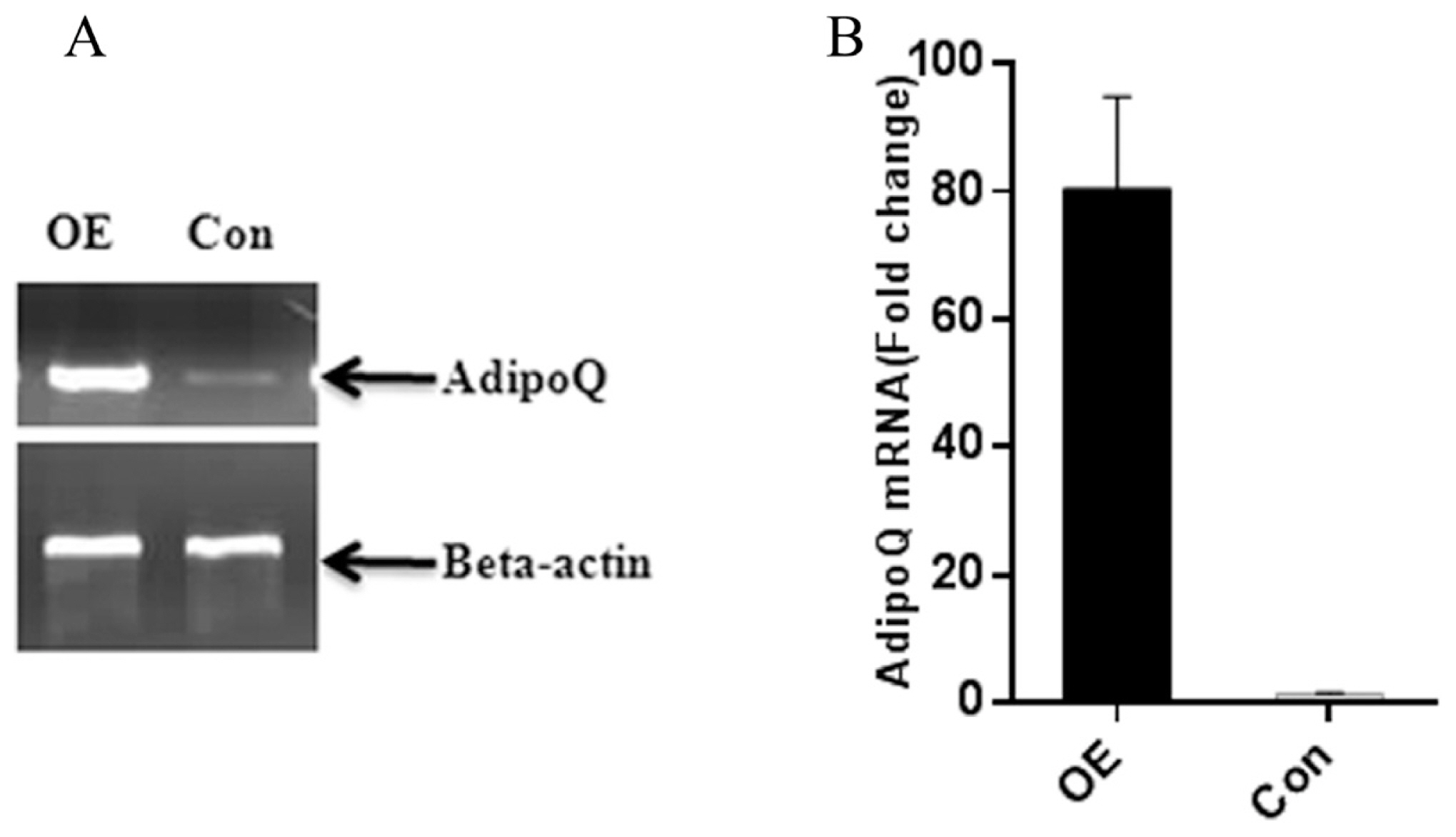
To determine the role of adiponectin in the adipogenic differentiation of goat MSCs, cells were transfected with the pEGFP-N1-AD plasmid to overexpress the goat AdipoQ gene. To evaluate effective overexpression of adiponectin, whole-cell lysates were harvested for analysis of expression levels. In SMSCs, adiponectin was overexpressed approximately 80-fold compared with that in control cells transfected with the pEGFP-N1 vector (Figure 6). These results demonstrate that cells transfected with the pEGFP-N1-AD plasmid can overexpress adiponectin compared with the expression levels of control goat SMSCs expressing green fluorescent protein (GFP).
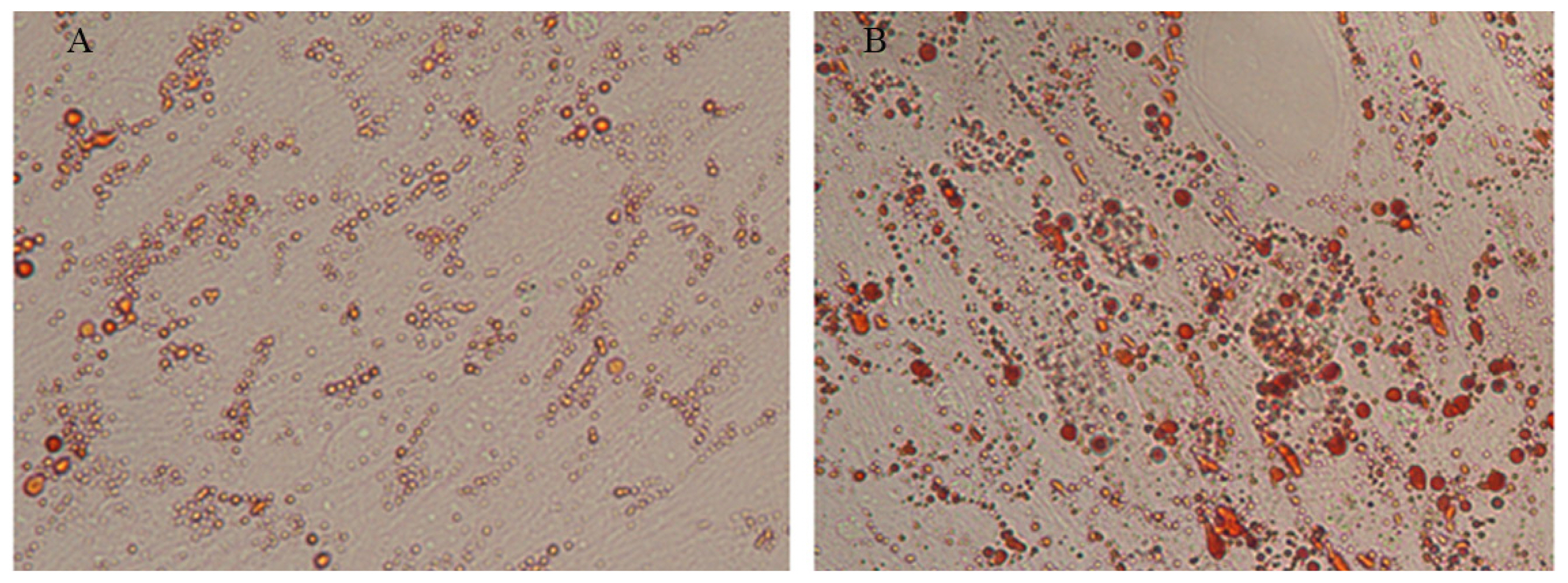
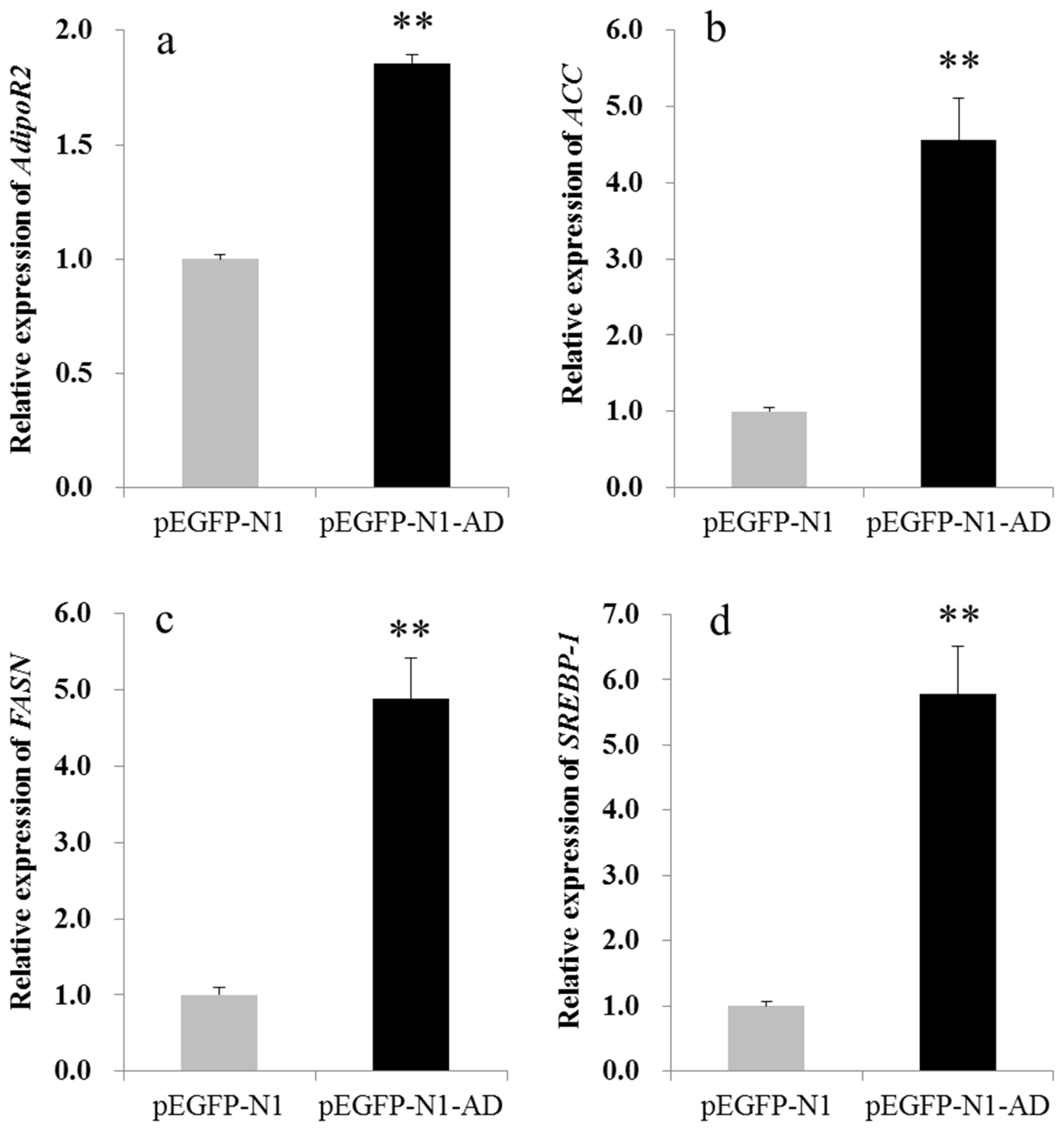
Next, satellite cells were induced with an MDI adipogenic cocktail and stained with oil red O seven days later. As shown in Figure 7, more oil red O-positive cells were observed in the pEGFP-N1-AD-transfected group than in the control group, indicating that adiponectin promoted MSC differentiate into adipocytes. In addition, several markers of adipogenic differentiation were detected by qPCR. The results showed that adiponectin overexpression significantly (p<0.01) up-regulated the mRNA expression levels of AdipoR2, ACC, FAS, and SREBP-1 (Figure 8), which were 1.85, 4.55, 4.88, and 4.25 times higher, respectively, than those of the control group. Conversely, no significant changes in expression were found for C/EBP╬▒, PPAR╬│, and AdipoR1 (data not shown).
DISCUSSION
Recently, many studies have focused on the functions of the AdipoQ gene in regulating glucose and fat metabolism and on its anti-inflammatory and anti-atherosclerosis effects in some species [12,22,23]. In the present study, we cloned the goat AdipoQ, AdipoR1, and AdipoR2 genes to investigate their expression and the functions of the encoded proteins. We found these gene sequences to be conserved among species. Adiponectin belongs to the complement factor C1Q family, and studies have proven that the C1Q domain has particular functions, including inducing SMC differentiation and fusion [24] and enhancing muscle fat oxidation and glucose transport [12]. The two receptors of adiponectin, AdipoR1 and AdipoR2, are seven-transmembrane domain proteins, but their protein structures lack homology with other mammalian proteins. In general, AdipoR1 and AdipoR2 are activated by different types of adiponectin. However, Narasimhan found that in yeast, the AdipoR1 and AdipoR2 proteins interact with osmotin, which has no sequence homology but does possess structural similarity to adiponectin [25]. This finding indicates that AdipoR1 and AdipoR2 might serve as receptors for other proteins.
Previous studies have shown that chicken AdipoQ is predominantly expressed in adipose tissue, whereas AdipoR1, and AdipoR2 are widely expressed in various tissues. AdipoR1 mRNA is highly expressed in skeletal muscles, adipose tissue and the diencephalon; AdipoR2 mRNA is most highly expressed in adipose tissue, followed by skeletal muscles and the liver [26]. Research in pigs showed that AdipoQ mRNA expression is abundant in adipose tissue, that AdipoR1 mRNA expression is abundant in skeletal muscle and adipose tissues, and that AdipoR2 mRNA is predominantly expressed in adipose and skeletal muscle tissues [27]. Overall, these three genes are mainly expressed in adipose and skeletal muscle tissues, where they are responsible for energy metabolism. Hence, the observed tissue distribution profile corresponds with their function in regulating lipid and fatty acid metabolism. In our study, the mRNA tissue distribution of AdipoQ and its two receptor genes was similar to that in chickens and pigs, suggesting a similar function among species.
A previous report stated that adiponectin expression and secretion are increased during the human adipocyte differentiation process, which is regarded as a distinctive marker of adipocyte differentiation [28]. Overexpression of adiponectin in 3T3-L1 fibroblasts increases the number and size of lipid droplets and promotes adipogenic differentiation [14]. Accordingly, these results suggest that adiponectin is able to promote fatty acid synthesis and adipocyte differentiation.
In this study, the adiponectin fusion protein was only found in the cytoplasm of SMSCs overexpressing the AdipoQ gene, indicating that adiponectin functions in the cytoplasm. Based on our qPCR results, the expression levels of AdipoR2, ACC, FASN, and SREBP-1 were significantly up-regulated, suggesting that adiponectin interacts with AdipoR2 to regulate expression of ACC, FASN, and SREBP-1 genes in SMSCs. It has been reported that ACC and FASN are two important factors for de novo fatty acid synthesis; ACC catalyzes the production of malonyl-CoA from acetyl-CoA, which is then converted into long-chain fatty acids by FASN [29]. Moreover, SREBP-1 is an important transcription factor that regulates ACC and FASN transcription for fat metabolism and adipogenic differentiation [30]. Thus, increased expression of these three adipogenic genes may result in fatty acid synthesis and lipid accumulation, which indicates that adiponectin may promote fatty acid synthesis and adipogenic differentiation.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print







