




INTRODUCTION
Exposure of feedlot animals to hot or cold stress reduces growth and feed efficiency (Ames et al., 1980). Cold climate during winter and exposure to cold stress induces metabolic acclimatization, resulting in decreased animal performance and production efficiency (Young, 1981; Birkelo et al., 1991). Temperature stress seemingly changes metabolic and endocrinal status of animals. For example, cattle exposed to acute cold have increased concentrations of plasma corticosteroids (Alvarez and Johnson, 1973) and circulating nonesterified fatty acid (NEFA) concentrations (Broucek et al., 1987; Nonnecke et al., 2009).
Low environmental temperature can cause cold stress, which affects the immune system and thus animal health. The effect of a cold environment on the immune response has been studied in other than cattle. For example, pigs in a cold environment caused increase in serum adrenocorticotropic hormone and cortisol concentrations coincident with tissue cytokine mRNA levels (Frank et al., 2003). Cold stress increases in vivo proinflammatory cytokine gene expression, including interleukin mRNA, in chicken (Hangalapura et al., 2005). However, limited data are available on the effect of a cold environment on the immune response in cattle.
Temperatures on the Korean peninsula increased by approximately 2┬░C during 1992 through 2004 due to a global warming (Ho et al., 2006), and hotter summers are expected in the future. In addition, global warming also caused colder winters in South Korea. Therefore, animal performance should have been greatly impacted by cold stress, as cattle sheds in Korea are generally of the open type. Little information is available on the effects of temperature on growth performance and the metabolic and immunological responses in Korean cattle steers. This study was performed to examine whether ambient temperature affects growth, feed efficiency, blood metabolites, and immune cell populations in Korean cattle steers.
MATERIALS AND METHODS
Animals and feeding trial
All experimental procedures involving animals were approved by the Seoul National University Institutional Animal Use and Care Committee (SNUIAUCC), Republic of Korea, and were conducted in accordance with the Animal Experimental Guidelines provided by SNUIAUCC.
Eighteen Korean cattle steers, with a mean age of 10 months and a mean weight of 277 kg, were used. The study was conducted at the University Animal Farm of the College of Agriculture & Life Sciences on the Pyeongchang campus of Seoul National University, South Korea. Experimental farm was covered by roof, and thus animal was protected from the rain. Doors were installed at both of barn, and animal was raised indoor. All steers were fed a growing stage- concentrate diet at approximately 1.5% of body weight using the DeLaval Alpro automatic feeding station (DeLaval, Tumba, Sweden) for 8 weeks. Feeding period 1 (P1) was from March 7 to April 3 and period 2 (P2) was from April 4 to May 1. The formula of concentrate diet is shown in Table 1. Timothy hay was fed ad libitum. Water was freely provided. The chemical composition of the feed is shown in Table 1. Daily feed intake of the concentrate diet was automatically recorded on an online Alpro system computer. An equal amount of hay was fed to each animal twice a day (8 AM and 6 PM), and the residual hay was weighed before the morning feeding. Samples of concentrate and hay were collected weekly and stored until analysis. Body weight was measured at 9 AM before feeding on the start date, and 4 and 8 weeks after feeding.
Chemical composition analyses
The chemical compositions (dry matter, crude protein, crude fat, ash, Ca, and P) of the concentrate diet and timothy hay were determined using an AOAC method (1990). The neutral detergent fiber and acid detergent fiber contents of the hay were analyzed using the sequential method with the ANKOM200 Fiber Analyzer (Ankom Technology Corp., Macedon, NY, USA) and reagents, as described by Van Soest et al. (1991).
Blood collection and temperature measurements
Blood was collected three times; on March 7, April 4, and May 2, 2014, after a 9 h fast by jugular venipuncture into non-heparinized vacutainers (20 mL; Becton-Dickinson, Waltham, MA, USA) and ethylenediaminetetraacetic acid (EDTA)-treated vacutainers (20 mL). Some of the whole blood collected in the EDTA-vacutainers was used for the immune cell analysis. Serum and plasma were separated by centrifugation at 1,500├Śg at 4┬░C for 15 min. The plasma and serum were subsequently stored at ŌłÆ80┬░C until analysis.
Ambient and climate temperatures and relative humidity inside and outside the barn were recorded at 1 h intervals using three HOBO data loggers (Onset Computer Corp., Bourne, MA, USA), and monthly average values of minimum, mean, and maximum temperatures and humidity were calculated using the daily data.
Blood analysis
The analytical reagents for glucose, triglyceride (TG), high-density lipoprotein (HDL), low-density lipoprotein (LDL), cholesterol, total protein, glutamic oxaloacetic transaminase (GOT), glutamic pyruvate transaminase (GPT), creatinine, blood urea nitrogen, calcium, magnesium, and phosphorus were purchased from JW Medical (Seoul, Korea). The analytical reagents for the NEFA, phospholipid, and beta-hydroxy butyrate analyses were purchased from Wako Pure Chemical (Osaka, Japan). The analytical reagents for complements C3 and C4 were purchased from Sekisui (Osaka, Japan). All of these parameters were analyzed using an automated chemistry analyzer (Hitachi 7180, Tokyo, Japan). Plasma cortisol was analyzed using a cortisol salivary HS enzyme-linked immunosorbent assay kit (cat. no. SLV4635; DRG Instruments, Marburg, Germany). The intra and interassay coefficients of variation for the cortisol kit were 4.0% and 4.6%, respectively, using bovine plasma samples.
Immune cell analysis
A 100 ╬╝L aliquot of whole blood was treated with 1 mL BD lysing buffer (Becton-Dickinson, USA) and incubated for 3 min at room temperature to analyze the immune cell characteristics. Total cell numbers were counted with a TC10 Automated Cell Counter (Bio-Rad, Hercules, CA, USA). Then, the granulocyte: lymphocyte ratio, identified by flow cytometry using the forward scatter (FSC) and side scatter (SSC) settings (Kampen et al., 2004), was calculated as a percentage of total cells. Leukocytes (5├Ś105 cells), stained with anti-bovine CD14-FITC and MHC class II DQ-PE antibodies for monocytes and B cells, respectively and CD4-Alexa647, CD8-PE, and CD25-FITC antibodies (AbD Serotec, Raleigh, NC, USA) for T cells, respectively, for 15 min at 4┬░C, were analyzed by flow cytometry (Becton-Dickinson, USA) and FlowJo software (TreeStar, Ashland, OR, USA). Briefly, lymphocytes and monocytes were gated based on FSC-SSC setting and CD14+MHC class II DQ+ cells were analyzed as monocytes and CD14ŌłÆMHC class II DQ+ as B cells. T cells were analyzed with CD4 and/or CD8 positive among lymphocytes and then activated T cells were identified as CD25+ cells.
Statistical analysis
All data are expressed as means┬▒standard error of mean. Differences in weather data among months were analyzed using one-way analysis of variance (ANOVA). Changes in the growth performance and blood parameters over time were analyzed by repeated-measures ANOVA with the TukeyŌĆōKramer post-hoc test. A p<0.05 was considered significant. All statistical tests were performed using R Studio for Windows software package (R Studio, Boston, MA, USA).
RESULTS AND DISCUSSION
Climate conditions
Experimental farm was covered by roof, and thus animal was protected from the rain. Doors were installed at both of barn, and animal was raised indoor. Thus, mild wind may affect wind-chill temperature. However, minimum indoor temperature was 1.0┬░C and 6.2┬░C on March and April, which are not within severe cold stress range. Thus, wind factor may be a minor climate variable. Mean (8.7┬░C) and minimum (1.0┬░C) indoor ambient temperature of March was lower (p<0.001) than that (13.0┬░C and 6.2┬░C) of April, respectively. But, maximum temperature of March (20.6┬░C) was not different from that of April (21.4┬░C). Mean (4.6┬░C), maximum (10.6┬░C), and minimum (ŌłÆ1.7┬░C) climate temperature of March was lower (p<0.001) than that (9.8┬░C, 15.8┬░C, 3.4┬░C) of April, respectively. Mean, maximum, and minimum relative humidity of March was not different from those of April (Table 2).
Growth performance
Daily intake of concentrate and forage was higher (p<0.001) during April (concentrate, 3.97 kg and forage, 3.71 kg) than March (concentrate, 3.54 kg and forage, 3.34 kg). Average daily gain was higher (p<0.001) during April (1.38 kg/d) than March (1.13 kg/d). Feed efficiency during April (0.18 kg gain/kg feed) was higher (p<0.02) than March (0.16 gain/kg feed) (Table 3). In this study, indoor ambient temperature affected both the growth and feed efficiency of Korean cattle steers. Weight gain and feed efficiency were lower during the colder month of March compared to April. Other studies have reported similar results; colder temperatures resulted in lower feed efficiency (Elam, 1970; Delfino and Mathison, 1991) and average daily gain (Birkelo et al., 1991). Cold stress has been categorized as ŌĆ£mildŌĆØ (0┬░C to ŌłÆ6.7┬░C), ŌĆ£moderateŌĆØ (ŌłÆ7.2┬░C to ŌłÆ13.9┬░C), and ŌĆ£severeŌĆØ (<ŌłÆ13.9┬░C) under dry winter cattle coat conditions (Grzych, 2010). Mader et al. (2010) suggested that animal susceptibility to stress varies more for cold stress than for heat stress dependent on breed, stage of animalŌĆÖs life cycle. Ambient temperatures well above the estimated lower critical value affect feedlot cattle growth performance (Webster, 1970; Young and Christopherson, 1974). Studies have suggested that energy requirements increased during winter or when animals are under cold stress due to the need to increase resting heat production to maintain body temperature by shivering or other thermogenic process (Young, 1983; Christopherson, 1985). Thus, the lower feed efficiency during March compared to April may have been due to the increased energy utilization for heat production at a colder ambient temperature. In results of our study, both climate condition and age effects may be compounded because climate effects were compared at different growing period. Generally, the younger animal has higher feed efficiency. But, feed efficiency was higher at period 2 in older age. Thus, climate condition may be a major effect, and age effect may be minor since only one month difference between 9.7 and 10.6 months of age.
Blood cortisol and metabolites
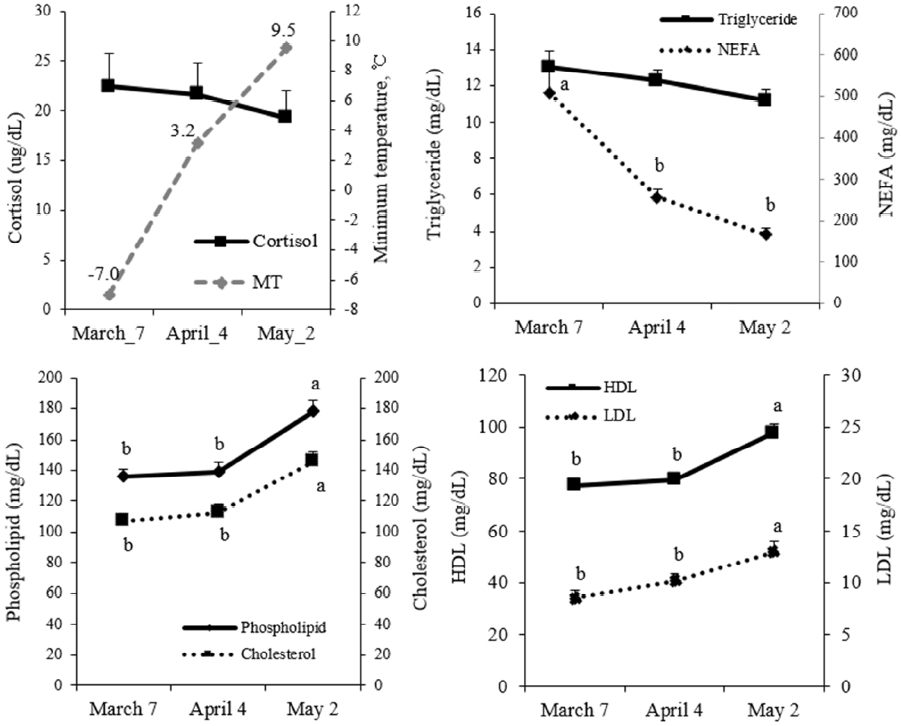
Blood NEFA concentrations were higher during the colder month of March than those during April and May (p<0.05; Figure 1), suggesting that lipolysis may occur at colder ambient temperatures to generate heat for maintaining body temperature. However, cortisol concentrations were not changed. These results suggest that the decrease in NEFA concentration may not be regulated by cortisol. Blood NEFA concentrations in dairy cattle often increase when feed intake cannot support their energy requirements, requiring the mobilization of NEFA by lipolysis of fat depots to support energy demand (Bauman and Currie, 1980). Broucek et al. (1987) reported that NEFA concentrations increase under low temperature conditions. Calves in a cold environment (4.7┬░C) also have higher NEFA concentrations than those in a warm indoor environment (15.5┬░C; Nonnecke et al., 2006). Taken together, increased NEFA concentrations during colder weather may help generate energy to maintain body temperature and growth. Stress is generally accompanied by an increase in cortisol concentration, and cortisol concentrations increase under cold conditions (Khan et al., 1970; Takahashi et al. 1986). In our study, the minimum temperature (1.0┬░C) during March may not have been sufficiently cold to stimulate cortisol secretion. Generally, amount of diet feed consumed significantly affects blood metabolites and hormone concentrations. Therefore, we collected blood after 9 h fasting to minimize such diet variation among animals. Mills and Jenny (1979) and Ward et al. (1992) reported that cortisol concentrations were increased during fasting. Thus, cortisol concentrations at fasting may be higher than that at feeding stage. Our study showed no significant difference in fasting cortisol concentrations between April and May. We cannot exclude that fasting condition masks temperature responses.
In the present study, blood concentrations of phospholipids, HDL, LDL, and cholesterol were lower during March and April than during May (p<0.05; Figure 2). In rat study, cold stress decreased serum total cholesterol and HDL concentrations (Tsopanakis and Tesserommatis, 1991). Consistent with our study, cold-stressed broilers have lower serum total cholesterol, HDL, and LDL concentrations (Chen et al., 2012). Collectively, our results suggest that ambient temperature may affect lipoprotein metabolism.
In our study, blood concentrations of glucose, TG, and total protein were not different (p>0.05) among months (Figures 1 and 2). Blood concentrations of urea nitrogen, GOT, GPT, phosphorus, and albumin were higher (p<0.05) during May than during March (Figure 2). Changes in these parameters due to cold stress or low temperature have not been reported previously.
Immune cell populations
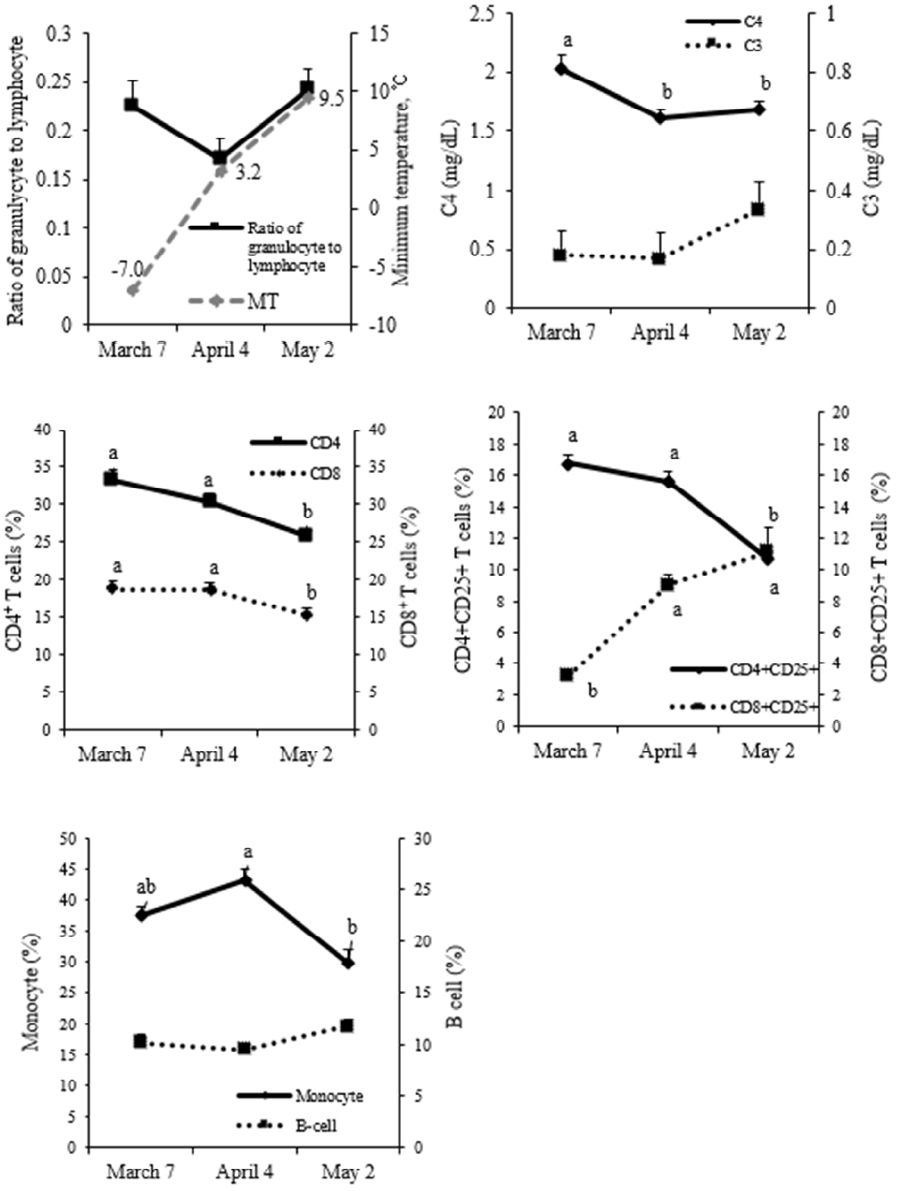
Inflammatory responses are often associated with the induction of acute-phase reactants, such as the C3 and C4 compartments of the complement system (Ghazavi et al., 2013). In our study, blood C4 levels were higher (p<0.05) during March than April or May, whereas C3 level did not differ (Figure 3). Our results suggest that the C4 complement factor may be induced more during the colder March temperatures than other months. The functional significance of this change remains to be elucidated.
The granulocyte: lymphocyte ratio and B cell and monocyte populations did not differ among months. It is well known that glucocorticoids released during stressful events often modulate the immune system, including increases in the neutrophil: lymphocyte ratio and changes in the number of monocytes (Weber et al., 2004). Neutrophil: lymphocyte ratio changes occurred under acute and inflammatory conditions, such as weaning stress and tissue injury (Kim et al., 2011). It is probable that the ambient temperatures during March in the present study were not low enough to induce such changes.
The numbers of CD4+, CD8+, and CD4+CD25+ T cells were higher (p<0.05), and the number of CD8+CD25+ T cells was lower (p<0.05), in March than in May (Figure 3). These results suggest that the number of helper T cells, cytotoxic T cells, and regulatory T cells are likely modulated during the colder month of March compared to May. T cells were increased significantly in the spleen of acute cold-stressed mice, whereas CD4+ and CD8+ cell populations were minimally increased (Kizaki et al., 1995). Whether these changes in blood T cell populations are directly associated with ambient temperature must be clarified in cattle.
In conclusion, the minimum ambient temperature was lower in March (1┬░C) than in April (6┬░C). Daily weight gain and feed efficiency were lower in March than in April, suggesting that ambient temperature affects growth and feed efficiency in Korean cattle steers. Higher circulating NEFA concentrations during March compared to April suggest that lipolysis may occur at colder temperatures to generate heat to maintain body temperature. Blood CD4+, CD8+ and CD4+CD25+ T cell populations were higher in March than in May, suggesting that ambient temperature affects blood T cell populations.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







