



INTRODUCTION
Racing horses, such as Thoroughbreds, have been selected for athletic performance traits by implementing systemic structural and functional adaptations. These processes have given racing horses a range of extreme physiological characteristics, including high aerobic and anaerobic metabolic capacities, a large lung volume, high maximum hemoglobin concentration and cardiac output as well as a large muscle mass to body ratio, high skeletal muscle mitochondrial density and oxidative enzyme activity, and large stores of energy substrates (Essén-Gustavsson and Lindholm, 1985; Pösö et al., 1993; Hyyppä, et al., 1997; Hinchcliff and Geor, 2008). Racing, as a sort of exercise, results in acute immune responses and inflammation in joints and muscle of horses (Auer et al., 1989; Bertone et al., 2001; Petersen et al., 2004; Firth et al., 2006). In horses, like human, intense physical activity induces subclinical tissue damage in muscle and joints, resulting in recruit of immune cells, i.e., neutrophils and monocytes. These cells trigger inflammation by up-regulating various cytokines, such as tumor necrosis factor alpha (TNFα), interleukin-1 beta (IL-1β), interleukin-6 (IL-6), and interleukin-10 (IL-10) (Streltsova et al., 2006; Lamprecht et al., 2008) and growth factors such as VEGFα in muscle (Breen et al., 1996; Roca et al., 1998; Richardson et al., 1999; Richardson et al., 2000; Brutsaert et al., 2002).
It is crucial for racing horses to recover from exercise as soon as possible. This recovery process might be mediated by soluble factors such as cytokines and growth factors. Nonetheless, there is no biomarker based on cytokines and/or growth factors that may reflect the degree of recovery from exercise in racing horses. In a previous study, we identified differentially expressed genes (DEG) in muscle and blood from Thoroughbred horses in response to exercise (Park et al., 2012). Furthermore, the findings provides a chance to develop molecular markers that may be related to economic traits of racing horses. The identified DEGs can be used to develop biomarkers that might be useful for monitoring the health of a horse after racing. We found that VEGFα expression was significantly increased in muscle, indicating that VEGFα may play an important role in recovery from exercise. In spite of the importance of VEGFα in various biological processes, such as angiogenesis, so far few studies have been conducted on this gene at the molecular level in horses. Therefore, in this study, we investigated the molecular structure of horse VEGFα gene, constructed a phylogenetic tree, and examined the expression profiles in various horse tissue samples. In addition, since there is a lack of suitable biomarkers to monitor recovery in horses after exercise, we explored the possibility of using VEGFα as a biomarker for exercise and recovery in blood leukocytes.
MATERIALS AND METHODS
Blood and tissue sample in horse
Blood samples were obtained from three male Thoroughbred horses (2 through 4 year-old; two horses: 2 year-old, one horse: 4 year-old) maintained at Ham-An racing Horse Resort and Training Center. Horses were placed on a treadmill and the treadmill exercises were performed at the speed of 10 to 15 km/h. The blood samples were taken before exercise and every 30 min after exercise up to 120 min. The National Institute of Subtropical Agriculture, Rural Development Administration, provided three male Jeju horses (4 year-old) that were used for tissue sampling. Skeletal muscle, kidney, thyroid, lung, appendix, colon, spinal cord and heart tissues were obtained from each horse and kept in liquid N2 until RNA extraction.
Sequence data analysis
The VEGFα DNA and amino acid sequences of ten species (cattle, chicken, chimpanzee, dog, horse, human, pig, mouse, rat, sheep) were retrieved from Ensembl (http://www.ensembl.org/), The Ensembl ID for the sequences are as follows: horse- ENSECAG00000009402, human- ENSG00000112715, chimpanzee- ENSPTRG000 00001080, mouse-ENSMUSG00000023951, rat- ENS RNOG00000019598, cow- ENSBTAG00000005339, pig: ENSSSCG00000001695, chicken- ENSGALG00000010290, dog- ENSCAFG00000001938, cat- ENSFCAG00000026462.
The retrieved sequences were aligned in BioEdit with the CLUSTERW option. Phylogenetic analysis was conducted using MEGA version 5.0 (Arizona State University, AZ, USA) (Tamura et al., 2011) The Neighbor Joining method (Saitou et al., 1987) was used with the following options: pairwise-deletion, 1,000 bootstrap replications and Kimura 2-parameter. Pairwise deletion was chosen to retain all sites initially, then excluding them as necessary in the pairwise distance estimation. Substitutions of nucleotides were obtained by using the Kimura 2-parameter model. We also showed that synonymous substitutions per site (Ks) and non-synonymous substitutions per site (Ka) in VEGFα genes for individual species. In addition, pairwise distance was calculated by analyzing the nucleotide and amino acid similarity.
RNA extraction and cDNA synthesis
Trizol reagent (Invitrogen, Carlsbad, CA, USA) was used to extract total RNA from the tissue samples (skeletal muscle, kidney, thyroid, lung, appendix, colon, spinal cord, and heart) and leukocytes after exercise according to the Invitrogen manual. In order to prevent contamination of genomic DNA, RNase-free DNase kit (Qiagen) was used according to the manufacturer’s manual. Total RNA quantification was performed using a NanoDrop ND-1000 Spectrophotometer (Thermo Scientific, Wilmington, DE, USA). The cDNAs were synthesized in a reaction with oligo-dT primers, moloney-murine leukemia virus (MMLV) reverse transcriptase (Promega, Madison, WI, USA), RNase inhibitor (Promega) and RNase-free ddH2O, which was incubated at 37°C for 4 h.
RT-PCR analysis of equine VEGFα
The horse VEGFα expression level was analyzed by reverse transcription-polymerase chain reaction (RT-PCR) amplification. The primers were designed using the PRIMER3 software (MIT, Cambridge, MA, USA, http://frodo.wi.mit.edu/primer3/) with an expected product size of 184 bp. To detect equine VEGFα mRNA, following primers were used: forward 5′-CTA CCT CCA CCA TGC CAA GT-3′ and reverse 5′-CAC ACA GGA TGG CTT GAA GA-3′. The RT-PCR conditions were as follows: an initial step of 94°C for 10 min, 35 cycles of 94°C for 30 s, 60°C for 30 s, 72°C for 30 s, and a final step of 72°C for 10 min. RT-PCR products were analyzed with gel electrophoresis on a 2.0% SeaKem LE agarose gel (Lonza, NJ, USA). Equine GAPDH gene was used as a normalizer for RT-PCR.
Quantitative PCR analysis of equine VEGFα expression and statistical analysis
The cDNA was analyzed by quantitative-PCR (qPCR) on BioRad CFX-96 (Bio-Rad, Hercules, CA, USA). Each reaction was performed in a total volume of 25μL with 14 μL SYBR green master mix, 2 μL (5 pmol) VEGFα forward primer, 2 μL (5 pmol) reverse primer, 5 μL distilled water and 2μL (50 ng/μL) of the cDNA. The PCR conditions were as follows: pre-denaturation step at 94°C for 3 min, 39 cycles at 94°C for 10 s and at 60°C for 30 s, 72°C for 30 s, followed by 72°C for 10 min as a final step. Dissociation was accomplished in a condition at temperature increase from 55°C to 95°C over 25 min. Dissociation temperature increased in 0.5°C each. All samples were measured in triplicate to ensure reproducibility, and Ct value was calculated using the 2−ΔΔCt method (Livak et al., 2001). Statistical significance was calculated by t-test. A p values of less than 0.05 were considered to indicate statistical significance.
RESULTS AND DISCUSSION
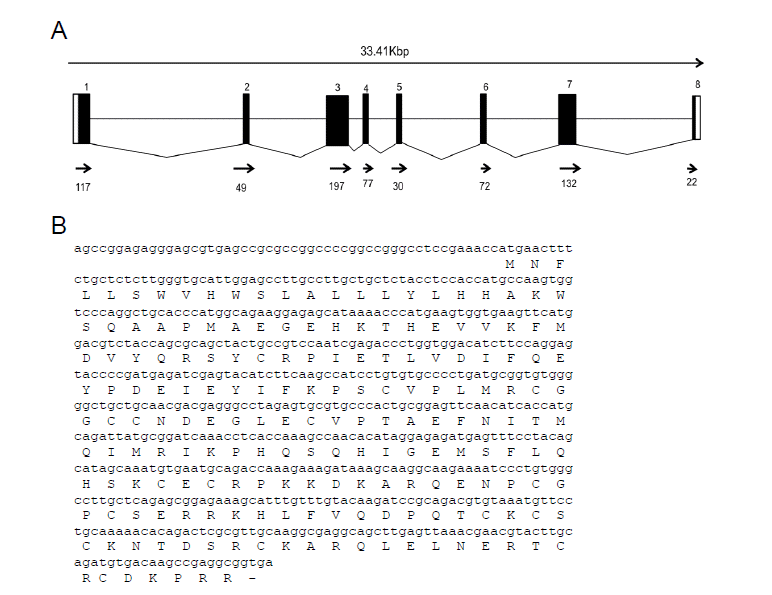
Molecular structure and distribution of the equine VEGFα gene
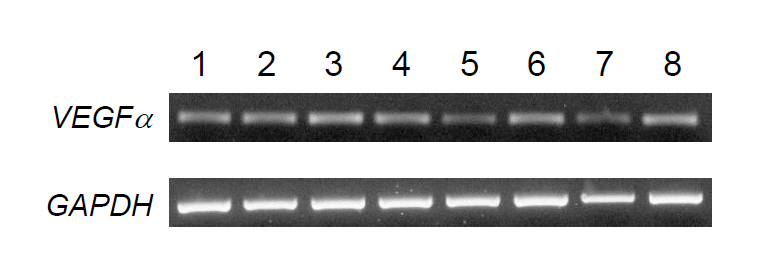
In a previous study, we performed RNA-seq analyses to identify DEGs in response to exercise. Among the identified DEGs, we found that the transcript of VEGFα was expressed up to 2.59 folds higher after exercise (Park et al., 2012). As not much is known about the horse VEGFα gene, we retrieved horse VEGFα sequences from the Ensembl database and analyzed its molecular structure. We found that the gene had eight exons and was 696 bp long in horses (Figure 1A). When we examined the expression of VEGFα transcripts by RT-PCR in eight tissues, we found that all tissues ubiquitously expressed VEGFα transcripts. However, thyroid and lung tissue expressed the highest levels of VEGFα transcript whereas the skeletal muscle and appendix tissue expressed relatively low levels of VEGFα transcript (Figure 3).
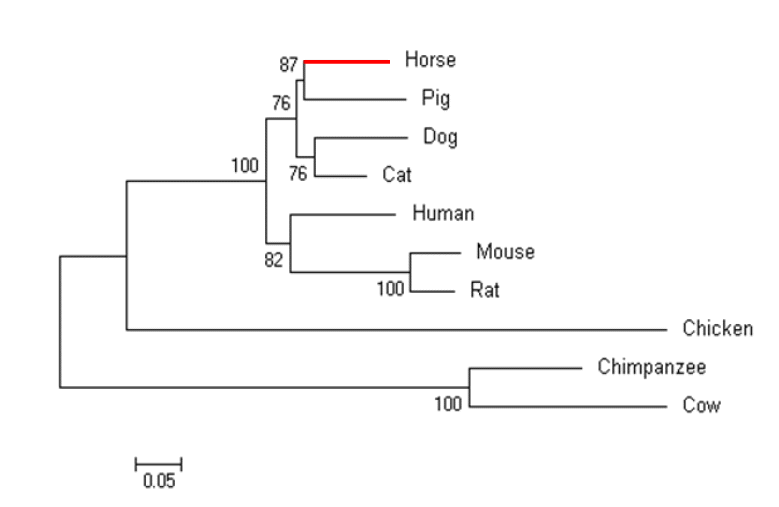
Evolutionary and phylogenetic analysis of VEGFα gene
Phylogenetic analysis of the VEGFα gene by neighbor-joining method revealed that equine VEGFα belongs to the same clade as the porcine VEGFα gene (Figure 2).
To better understand the evolution of the VEGFα gene, we analyzed synonymous (Ks) and non-synonymous (Ka) substitutions of VEGFα amino acids from ten species (Table 1). Pairwise comparison showed that Ks ranged from 0.168 to 2.112, whereas Ka ranged from 0.191 to 1.468. It is noted that Ks of equine VEGFα was the lowest with the pig gene (0.188) and the highest with the dog gene (2.112). The lowest Ks values were obtained between mouse and chimpanzee (0.092). The average of Ka and Ks were 0.839 and 0.689, respectively, resulting in a Ka/Ks ratio of greater than 1. This means that non-synonymous substitutions occurred more rapidly than synonymous substitutions. These results are based on the assumption that the protein-coding regions in the human genome have been under positive selection during evolution (Bustamante et al., 2005). These data indicated that positive selection occurred in the VEGFα genes during the evolutionary history of the horse.
Differential expression of equine VEGFα transcripts during recovery post exercise
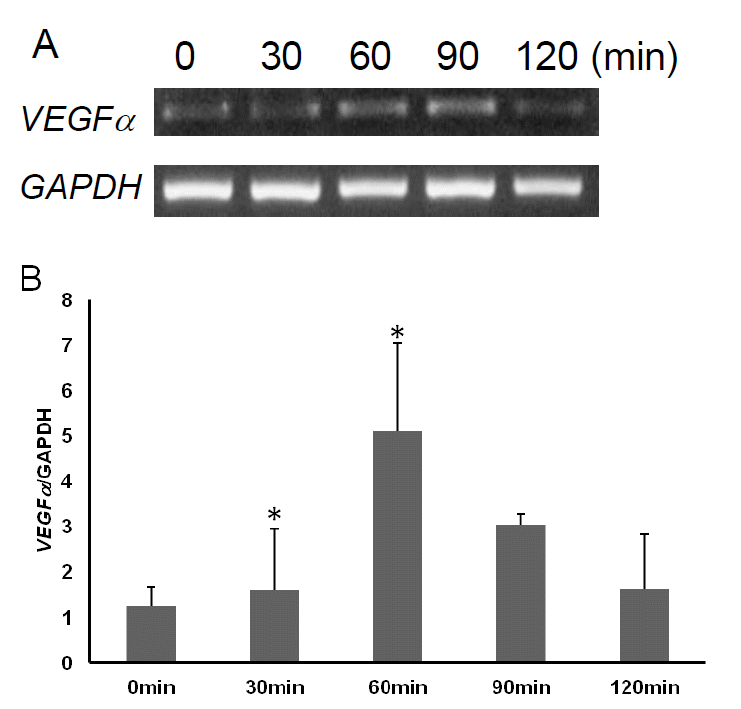
To investigate the relationship between VEGFα gene expression level and time length after exercise, we performed qPCR analysis using blood leukocyte samples obtained before and after exercise (30 min, 60 min, 90 min, 120 min) from three horses (Figure 4A and B). Interestingly, the expression level of VEGFα was the highest after 60 min of exercise and then gradually decreased after upto 120 min. But the expression level of 120 min was slightly higher than that of 0 min (before exercise, p>0.05).
CONCLUSION
In horse, as there are few reliable biomarkers for recovery after exercise except creatine kinase (Linder et al., 2006) and lactate (Lindner et al., 2009), it is necessary to identify and evaluate markers for post exercise recovery. In this sense, RNA-seq based transcriptome analysis may be useful. In a previous study, horse VEGFα transcript increased significantly in response to exercise (Park et al., 2012) similar to humans suggesting a conserved mechanism for the regulation of VEGFα transcription. Therefore, we selected the equine VEGFα gene for further evaluation at the molecular level. A phylogenetic analysis showed that the horse VEGFα gene is grouped into the same clade as the pig gene and have undergone positive selection. In terms of expression, horse VEGFα was found to be ubiquitously expressed but regulated by exercise in blood leukocytes. Especially, expression kinetics obtained in this study showed that the transcript level gradual increased up to 60 min (p<0.05), and then remained at a slightly higher level than at time 0, which is not statistically significant (p>0.05). It is not certain how VEGFα expression is upregulated and reached the highest level after 60 min of exercise. One explanation would be that acute exercise may induce the hypoxia condition in leukocytes, which triggers hypoxia-inducible factor-1α (HIF-1α) activation. Activated HIF-1α will induce the transcriptional activation of VEGFα by binding to HIF-responsive elements which are located in the promoter region of VEGFα (Jiang et al., 1996). Therefore, it is worth examining equine HIF-1α expression in blood leukocytes after exercise to reveal the causal relationship between HIF-1α and VEGFα expression.
Nonetheless, the role of VEGFα in response to exercise and recovery is not clear in blood. It is likely that the circulating VEGFα facilitates mobilization of monocytes into injured areas after exercise, and promotes wound healing by stimulating endothelial cell proliferation and extracellular matrix changes as process of recovery (Rocha et al., 1988; Richardson et al., 1999). Additionally, vascular distribution and density increase in response to exercise in skeletal muscle as an adaptation, which accompanies increase VEGFα expression (Brodal et al., 1977). Further study is warranted to explore the relationship between expression levels of VEGFα in muscle and blood with physiological changes.
In summary, we analyzed molecular structure of equine VEGFα gene and performed phylogenetic analysis with homologues of other species, which revealed that VEGFα gene of horse belongs to the same clade with that of pig. Gene expression analysis using various horse tissues showed the ubiquitous expression of horse VEGFα gene. Also, it is of note that VEGFα expression in blood leukocytes is regulated in response to exercise, suggesting that horse VEGFα expression in leukocytes can be used as a biomarker for recovery after exercise.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







