



INTRODUCTION
Surimi, a stabilized myofibrillar protein from fish, is the primary ingredient in fish paste or imitation crab. It is prepared by separation, washing, and mincing to eliminate undesirable blood, lipids, enzymes, and sarcoplasmic proteins (Vilhelmsson, 1997; Park and Morrissey, 2000). In general, surimi is light in color, bland in odor, low in fat, and extremely functional due to the unique gelling properties of the myofibrillar proteins. These properties make surimi a robust functional ingredient for fabricating new food products (Han-Ching and Leinot, 1993; Lanier, 2000). Numerous studies have been conducted on surimi containing fish meat. In addition, application of surimi technology in the production of surimi-based products using the proteins from other animal species could provide a new approach for increasing its utilization and functional properties.
Animal meat is of much interest in the development of surimi-based products such as those made from beef, pork and chicken (Park et al., 1996; Srinivasan and Xiong, 1996; Kristinsson and Hultin, 2003). Pork leg meat, for instance, has been reported to have high myofibrillar protein content allowing increased gel forming capacity in surimi. Thus chicken or pork meat is an attractive substitute for fish meat surimi (Jin et al., 2007), with spent laying hens providing a particularly economical source of surimi-based products. Indeed, it was recently shown that spent laying hen meat has potential as filler in surimi-based products (Hur et al., 2011). Therefore, the addition of myofibrillar proteins from spent laying hens to surimi-based imitation fish paste (IFP) holds promise in promoting the utilization of such products.
It is important for the meat industry to develop new products to satisfy emerging consumer demands for high quality foods. Protein hydrolysates, rich in low molecular weight peptides (di- and tri-peptides, with minimal free amino acids), are a good dietary source due to their high nutritional value and therapeutic properties (Bhaskar et al., 2007). In recent years, research has focused on the generation of bioactive peptides from food sources including meat and meat by-products (Daoud et al., 2005; Li et al., 2007). Peptides have been shown to exert antioxidant, antimicrobial, and antihypertensive effects (Saiga et al., 2003a; Jang et al., 2008; Liu et al., 2009; Escudero et al., 2010). Some peptides possess antihypertensive activity through their ability to inhibit Angiotensin I-converting enzyme (ACE) (Arihara et al., 2001). Also, bioactive peptides can be used as components in functional foods. Protein hydrolysates have been shown to enhance the emulsifying and foaming properties of fish (Shahidi et al., 1995). These peptides need first to be released from the original protein during food processing or digestion in order to exhibit biological activity and little is known about this process in meat and meat products.
Mechanically deboned chicken meat (MDCM) has a high content of heme pigments, connective tissue, and fat (Yang and Froning, 1992). It is dark in color, has undesired textural properties, and is susceptible to lipid oxidation. However, MDCM is one of the most common raw materials used to produce processed poultry products. Also, MDCM is widely used in the food industry for upgrading the functional and nutritional properties of proteins, thereby creating value-added products. Thus, MDCM hydrolysis may be used as a potential starting material for the generation of bioactive peptides.
The aims of the present investigation were to study the effects of added MDCM hydrolysates on the gel properties, oxidative stability, and angiotensin I-converting enzymes (ACE) inhibitory activities of IFP made from Alaska Pollack and spend laying hen meat.
MATERIALS AMD METHODS
Sample preparation
Frozen Alaska Pollack were purchased from Han-sung Food Co. Ltd. (Pusan, Korea), cut into 500 g blocks while frozen, packed into polyethylene bags, and stored at −20°C until use. Spent laying hens also were obtained at the same time from a commercial slaughterhouse. Spent laying hen myofibrillar protein was collected by the pH adjustment (Jin et al., 2008). MDCM hydrolysates were obtained by the protein hydrolysates method. Three batches were collected on different days for experimental replication. IFP samples were divided into three groups: Control (C); composed of commercial IFP containing spent laying hen meat, T1; consisted of IFP sample containing 0.4% MDCM hydrolysate, and T2; consisted of IFP sample containing 0.8% MDCM hydrolysate. The composition of the IFP is presented in Table 1 and a flow diagram depicting IFP preparation is shown in Figure 1.
pH method
The external fat tissue, bone, and skin were removed from the muscles, and the lean muscle was cut into approximately 3.0×3.0×2.0 cm3 cubes and ground through a 3 mm diameter hole using a mincer. The minced samples were combined with six times volume of distilled water and homogenized with a Polytron homogenizer (T25-B, IKA Sdn. Bhd., Malaysia) at 8,000 rpm for 30 s. The pH of the homogenate was then adjusted to pH 11.0 by the addition of 1 mol NaOH and centrifuged at 10,000×g for 25 min, after which the top layer of fat and bottom layer of connective tissue were discarded. The middle layer containing the myofibrillar protein was then adjusted to pH 5 by addition of 1 N HCl solution, after which it was centrifuged at 10,000×g for 25 min. The resulting sediment was then used for the manufacture of IFP.
Preparation of the protein hydrolysates
In this study, one- and two-stage hydrolysis was employed. The nerves, skin, and visible fat were removed from the meat, which was then fragmented, ground, and homogenized with distilled water (meat:water ratio, 1:3 w/w). The homogenate was heated at 43°C and the pH was adjusted to 7.0 with 2 N NaOH. Five percent of Protarmex) was added and the reaction pH was maintained at a constant pH by the addition of 2 N NaOH. The hydrolytic process was terminated by heating the mixture at 85°C for 20 min, ensuring inactivation of the enzyme. The resulting slurry was centrifuged (Union 5KR, Hanil, Gangneung, Korea) at 8,000 rpm for 10 min to remove insoluble fractions. The hydrolysate slurry was then heated to 50°C and the pH was adjusted to 7.0 with 2 N NaOH. Bromelain (1%) was added to the mixture and the reaction pH was maintained by the addition of 2 N NaOH. After heating at 90°C for 15 min to inactivate the enzyme, the hydrolysate was centrifuged at 8,000 rpm for 20 min to remove insoluble fractions. The degree of hydrolysis (DH) was determined using the 20% (w/v) trichloroactic acid (TCA) method, as described in the semi-micro Kjeldahl procedure. DH was defined as the percentage ratio of the total nitrogen in two-stage hydrolysate (A) to the total nitrogen in one-stage hydrolysate (B), and calculated as ([A–B]/B)× 100 (4.36% crude protein and 24.7% DH). The protein hydrolysate was then stored at 4°C until use.
Proximate composition
The proximate composition analysis of IFP batters including moisture, crude protein, crude fat, and crude ash, was performed according to AOAC methods 950.46, 992.15, 985.15, and 920.153 for sausage batter (AOAC, 2000). Moisture, protein, fat, and ash parameters were determined in triplicate from IFP product.
Gel characteristics
The gel characteristics of cooked IFP samples were determined according to the method described by Phatcharat et al. (2006). Three cylindrical pieces 3.5 cm wide and 3 cm thick were maintained at 20°C prior to measuring. The breaking force, deformation, gel strength and jelly strength were measured using a texture analyzer (EZ-test, Shimadzu, Tokyo, Japan) equipped with a cylindrical plunger (diameter 5 mm, depression speed 80 mm/min).
Water-holding capacity (WHC)
The water-holding capacity of IFP samples was determined by the method described by Hughes et al. (1997). Samples (approximately 10 g) were placed in 50 mL plastic centrifuge tubes and heated for 15 min in a water bath (90°C). The samples were then cooled to room temperature and centrifuged at 9,000×g at 4°C for 20 min (Union 5KR, Hanil, Korea). The supernatant was eliminated and the WHC of the remaining pellets were calculated as follows: WHC (%) = 1−([weight of sample before heating-weight of sample after heating and centrifugation]/total water content in the sample×100).
2,2-diphenyl-1-picryhydrazla hydrate (DPPH) radical scavenging activity
The DPPH radical scavenging activity measurement was modified according to the method of Bersuder et al. (1998). 500 μL of each peptide fraction was mixed with 500 μL of ethanol and 250 μL of a DPPH solution (0.5 mM 1,1-diphenyl-2-picrylhydrazyl/ethanol). The mixtures were incubated for 30 min in the dark at room temperature and the reduction of DPPH radicals was measured at 517 nm. DPPH radical scavenging activity was calculated as: DPPH radical scavenging activity (%) = ([absorption of control − absorption of sample]/absorption of control)×100. The control was conducted in the same manner, with the exception that distilled water was used instead of sample.
Lipid oxidation
Lipid oxidation was determined using the thiobarbituric acid reactive substances (TBARS) method (Buege and Aust, 1978). Cooked IFP sample (5 g) was weighed into a 50 mL test tube and homogenized with 15 mL of deionized distilled water using the Polytron homogenizer for 15 s at the highest speed (T25basic, IKA, Selangor, Malaysia). The IFP sample homogenate (2 mL) was transferred to a disposable test tube (13×100 mm) and butylated hydroxyanisole (10%, 50 μL) and thiobarbituric acid/trichloroacetic acid (TBA/TCA) solutions (4 mL) were added. The sample was mixed using a vortex mixer, incubated in a boiling water bath for 15 min to allow color development, and cooled at room temperature. Absorbance was determined at 531 nm against a blank containing 2 mL of deionized distilled water and 4 mL of TBA/TCA solution. The TBARS measure was expressed in mg of malondialdehyde (MDA) per kg of sample.
Angiotensin I-converting enzyme (ACE) inhibitory activity
ACE activity assay
The activity of ACE was determined using hippuryl-His-Leu (HHL) with the methods reported by Janitha et al. (2002). The assay was conducted in a Borate buffer (0.1 M, pH 8.3). The assay volume consisted of 150 μL of the ACE enzyme solution and 40 μL of assay sample. All of the solutions were incubated for 10 min at 37°C. 5 mL of HHL (0.3 M) and 100 μL of 0.1 M Borate buffer (pH 8.3) were added and incubated for 30 min. 1 M HCl (150 μL) was added to stop ACE activity. The reaction mixture obtained was used to quantitate the hippuric acid produced due to ACE activity on the substrate.
HPLC determination of hippuric acid content
A reversed-phase C18 column (Bonclone C18, 10 μM, 50×1.0 mm) protected by a guard column (Bondclone C18, 5 μM, 250×4.6 mm, Phenomenex Co., Torrance, CA, USA) was employed. The injection volume used was 20 μL. Elution of hippuric acid was detected by monitoring the absorbance at 228 nm. The control reaction mixture contained 40 μL of buffer instead of the assay sample. The percent inhibition of enzyme activity was calculated as follows: ACE inhibition (%) = ([hippuric acid of control-hippuric acid of sample]/hippuric acid of control)×100. The concentration of hydrolysate needed to inhibit 50% of ACE activity was defined as the IC50 value.
Statistical analysis
The study was designed as a 3×4 factorial experiment with treatment (control, T1 and T2) and storage time (0, 2, 4, and 6 weeks). The experiment was replicated three times. Data were analyzed using the general linear model (GLM) (SAS, 2000). Duncan’s multiple test was used to determine the statistical significance among the means at a 95% significance level.
RESULTS AND DISCUSSION
Proximate composition
The proximate composition of IFP with added MDCM hydrolysates is shown in Table 2. In this study, the IFP showed no significant difference in crude protein compared to the control during storage. However, all treatments with MDCM hydrolysates had lower moisture content compared to control at 0 and 6 weeks storage periods (p<0.05). Crude fat content in the MDCM hydrolysate-added groups was significantly higher than in the control at week 0 of storage. Likewise, the ash content in the 0.8% MDCM hydrolysate-added sample was significantly lower than in the other groups at 6 weeks of storage (p<0.05). The functional and textural properties of surimi depend on many factors including thermal conditions and various gelling and non-gelling ingredients (Nowasd et al., 2000). In general, high protein, low fat, and adequate water are required to make high quality surimi-based products (Jin et al., 2007), Moisture and fat are critical factors in surimi products (Uddin et al., 2006) and excessive lipids may adversely affect quality, due to oxidized lipids interacting with proteins (Smith, 1987). Also, protein concentration greatly affects the gel properties (Luo et al., 2004). However, we assumed that the protein contents would not influence the physical properties of IFP in this study because protein showed no consistent trends among the samples.
Gel characteristics
Gel characteristics of IFP with added MDCM hydrolysates are shown in Table 3. The breaking force values increased during storage in T1 and T2. The breaking force was lower in T2 as compared to T1 until 2 weeks storage (p<0.05). The gel strength and jelly strength showed consistent trends among the IFP samples; however, treatment with MDCM hydrolysates imparted higher breaking force values than control group at 4 and 6 weeks. Also, deformation, gel strength, and jelly strength were improved with added MDCM compared to the control group at 4 or 6 weeks (p<0.05). Protein concentration has a major positive effect on the breaking force (Luo et al., 2008). It has also been reported (Sylvia et al., 1994) that the gel-forming ability of surimi increases with decreasing water content, a result of higher myofibril protein concentrations and increased cross-link density. Therefore, our results indicate that the gel characteristics are favorably influenced by changes of proximate compositions and the addition of MDCM to IFP.
Water holding capacity (WHC), DPPH radical scavenging activity and lipid oxidation
Water holding capacity (WHC), DPPH radical scavenging activity, and lipid oxidation of IFP with added MDCM hydrolysates are shown in Table 4. The WHC was lower when MDCM hydrolysates were added compared to the control group during storage (p<0.05). During cooking, various meat proteins denature causing structural changes, shrinkage of meat fibers, and gel formation of myofibrillar proteins (Tornberg, 2005). Therefore, our current results suggest that the addition of MDCM to IFP may not directly affect WHC, but that with extensive degradation of muscle fibers an apparent WHC reduction occurs.
All treatment samples demonstrated increased DPPH radical scavenging activity during storage, while IFP showed significantly decreased DPPH radical scavenging activity after 2 weeks (p<0.05). DPPH radical scavenging activity was higher in the MDCM hydrolysate treatment groups compared to the control up until 4 weeks. The free radical scavenging activity and antioxidant activity; including the ability to donate hydrogen, to stabilize or terminate radicals, to sequester pro-oxidative metal ions, and to form a physical barrier around fat droplets, is determined by a specific amino acid composition and sequence (Kong and Xiong, 2006). Additionally, hydrolysate antioxidant activity depends upon the enzyme used. It has been reported (Wu et al., 2003) that DPPH activity improved for mackerel protein hydrolysate generated by Protease N. Another group (Wu et al., 2004) found that porcine hemoglobin hydrolysates, prepared through hydrolysis by Alcalase followed by Flavourzyme, exhibited high ferrous ion chelating abilities and DPPH radical scavenging activity. In our research, the DPPH radical scavenging activities of the Alcalase hydrolysates were similar to those reported by Wu et al. (2004).
The TBARS value increased in correlation with increasing storage periods in all IFP samples. In the treatment groups, TBARS values rapidly increased at 2 weeks (p<0.05) while they did not differ significantly among IFP samples until 4 weeks. TBARS values were higher in the MDCM hydrolysate treatment groups compared to the control at 6 weeks (p<0.05). Ahn et al. (1993) reported that differences in fat content, fatty acid composition, and the classes of lipids present effected the lipid oxidation of stored turkey patties. Meat products with a high degree of unsaturation are more susceptible to lipid oxidation (Morrissey et al., 1998). Accordingly, lipid oxidation in the present study was influenced by fat content (Table 2). Previously, Sakanaka and Tachibana (2006) reported reduction of TBARS values in beef homogenate by egg-yolk protein hydrolysates. However, hydrolysates possess a lower antioxidant activity in meatballs (Flaczyk et al., 2006). Hydrolysates at doses ranging from 1% to 2% slowed lipid oxidation of pork patties but their activity remained weaker than synthetic antioxidants (Pena-Ramos and Xiong, 2003).
Angiotensin I-converting enzyme (ACE) inhibitory activity
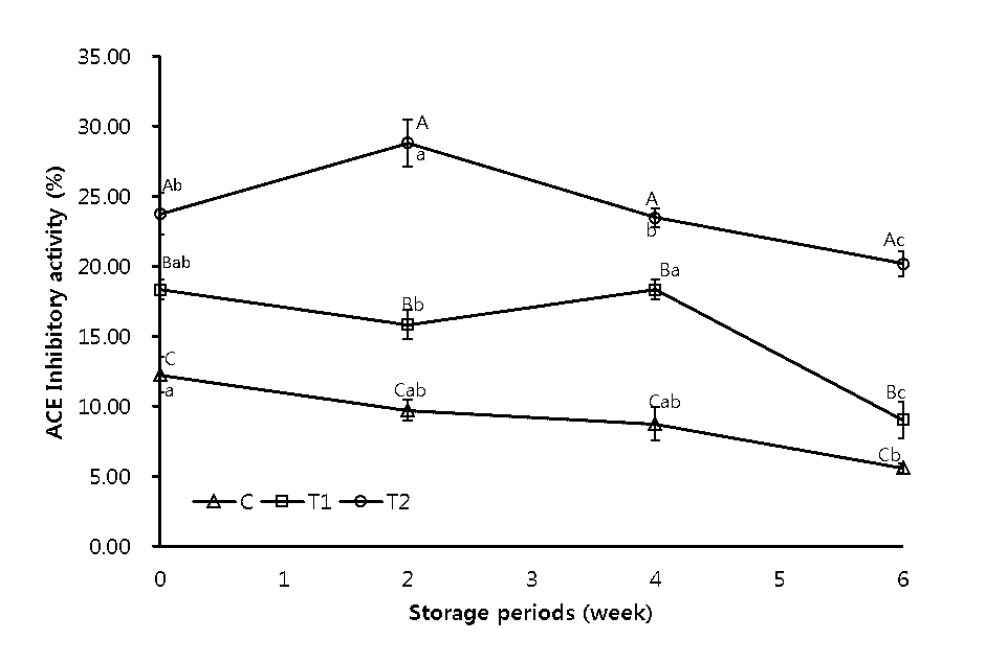
The ACE inhibitory activity in IFP with added MDCM hydrolysates is shown in Figure 2. ACE inhibitory activity of IFP showed a decreasing trend with increasing storage time. However, the ACE activity in the IFP with added treatments was lower than that of control. Addition of 0.8% MDCM hydrolysates was found to be more effective at inhibiting ACE activity of IFP compared to addition of 0.4% MDCM hydrolysates (p<0.05). ACE inhibitory peptides have been discovered in various animal sources such as porcine and chicken muscle. Specific ACE inhibitory peptides, Met-Asn-Pro-Asn (IC50 = 66.6 μM), Asn-Pro-Pro (IC50 = 290.5 μM), and Thr-Asn-Pro (IC50= 207.4 μM) were discovered in a porcine myosin hydrolysate (Arihara et al., 2001). Gly-Phe-Hyp-Gly-Thr-Hyp-Gly-Leu-Hyp-Gly-Phe (IC50 = 42 μM) was isolated from chicken breast muscle hydrolysate (Saiga et al., 2003b). Ahhmed et al. (2009) reported that mixing 5% meat hydrolysate from the porcine muscle biceps femoris with normal diet in rats resulted in clear positive effects on a common lifestyle-related disease such as hypertension. ACE plays an important role in the regulation of blood pressure as well as fluid and salt balance in mammals. Therefore, our results support the conclusion that MDCM hydrolysates can be used as a good source of health-promoting constituents in functional foods.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







