





INTRODUCTION
Cashmere is a precious natural protein fiber that derives from the secondary hair follicles (SHFs) of cashmere goats, and it has been regarded as high-grade textile raw material [1]. As one of the main products from cashmere goats, cashmere has important economic implications to local farmers and herdsmen. The morphogenesis and growth of cashmere is controlled by the activity of SHFs in cashmere goats [2]. Under the dermal papilla cell (DPC)-derived signals, the induced activation of SHF stem cells is significantly important for the reconstruction of SHFs with the subsequent the morphogenesis and growth of cashmere fibers in cashmere goats [3]. It is known that the induced activation of hair follicle stem cells is coordinately regulated by a variety of endogenous regulatory factors [4]. There is evidence that the Wnt/β-catenin signals can induce the activation of hair follicle stem cells [5]. There are also other signals and molecules implicated in the induced activation process of hair follicle stem cells, such as mammalian target of rapamycin [6], tumor necrosis factor [7], platelet-derived growth factor subunit A [8], and Foxi3 [5]. Also, it was reported that some miRNAs were also implicated in the activation event of hair follicle stem cells [4], such as miR-339-5p [9] and miR-214 [10].
N6-methyladenosine (m6A) has been revealed to be the most abundant modification in linear RNA molecules. Over the past few years, extensive m6A modifications sites are also found in many circRNA molecules that play significant roles in a variety of biological cells through m6A modification dependent model [11]. To date, a considerable number circRNAs have been identified in skin tissue or SHFs of cashmere goat [12], and some of them are demonstrated to positively regulate the differentiation of SHF stem cells into hair follicle lineage, such as circRNA-1926 [13], circRNA-1967 [14], and circRNA-0100 [15]. Moreover, some circRNAs in SHFs of cashmere goats are verified to contain multiple m6A modification sites [2]. Increasing lines of evidence indicate that the m6A modifications are heavily implicated in the functional exertion of the m6A modified circRNAs (m6A-circRNAs) via m6A-dependent model [11, 16].
In our recent study, a novel m6A-circRNA, named as m6A-circRNA-ZNF638, was identified in SHFs of cashmere goats, and it was transcribed from the antisense strand of goat zinc finger protein 638 (ZNF638) gene with significantly higher expression at anagen SHFs than that at telogen [2]. As a matter of fact, the ZNF638 is a member of ZNF family. Although the functional roles of ZNF638 in SHFs of cashmere goat remains unclear, several members of this family (ZNFs) were demonstrated to play significant roles in the physiological processes of cashmere goat SHFs including g ZNF264, ZNF347, ZNF407, ZNF454, ZNF667, and ZNF704 [17]. Considering that the SHF stem cells are under the vigorous status of induced activation at anagen, we hypothesize that the m6A-circRNA-ZNF638, as a circular RNA molecule transcribed from the antisense strand of ZNF638 gene, may be implicated in the induced activation process of SHF stem cells of cashmere goats.
In this study, we firstly assessed the relative expression of m6A-circRNA-ZNF638 in SHF bulge of cashmere goats at anagen and telogen. Further, we evaluated the putative effects of m6A-circRNA-ZNF638 on the induced activation of SHF stem cells of cashmere goats and investigated its possible functional mechanisms. Our results provided new molecular evidence for unveiling the potential molecular mechanisms of the induced activation event of SHF stem cells in cashmere goats.
MATERIALS AND METHODS
Sample collection, preparation, and total RNA isolation
In this study, all experimental protocols were reviewed and approved by the Experimental Animal Ethics and Welfare Committee of Shenyang Agricultural University (Shenyang, China) under the ethical code: 201606005. Correspondingly, the experiments were performed based on the approved procedure guidelines. The skin tissues (body side) were collected from nine cashmere goats (Liaoning cashmere breed, female, three-year-old, no traceable common genetic relationships) as described in our previous investigation [14]. In brief, the skin tissues were sampled from each goat using a sterilized surgical blade. The collected skin samples were cleaned with 75% alcohol. Subsequently, the skin samples were cut into pieces of 5 mm2 and washed with PBS for 3 times. The samples were further digested at 4°C with dispase II (0.25%; Roche, Mannheim, Germany) for 8 h. Under a stereo microscope, the SHFs were isolated with a sterilized scalpel and disposable syringe needle, and the SHF bulge sections were isolated following described protocol by Ohyama and Kobayashi [18]. The total RNA was extracted from SHF bulge using RNAiso reagent kit (TaKaRa, Dalian, China) according to the manufacturer’s recommendations.
Sequence of m6A-circRNA-ZNF638 and its in-silico analysis
The analyzed m6A-circRNA-ZNF638 has been identified from SHFs of cashmere goats in our previous study [2]. The sequence of m6A-circRNA-ZNF638 was managed and displayed with BioEdit software [19]. We determined the transcribed source of m6A-circRNA-ZNF638 in goat genome through aligning its linearized sequence into the goat genome (https://www.ncbi.nlm.nih.gov/genome/?term= goat, assembly, ARS1, last access: 18 March 2022). The potential target binding miRNAs within m6A-circRNA-ZNF638 sequence were predicted using two procedures (RNAhybrid: https://bibiserv.cebitec.uni-bielefeld.de/rnahybrid, and miRDB: http://www.mirdb.org, last access: 18 March 2022), and were further selected among the resulting miRNAs through taking an intersection. The goat miRNA sequences were retrieved from the miRNA database: miRNAsong (https://www2.med.muni.cz/histology/miRNAsong/index.php, last access: 18 March 2022). The potential m6A sites within circRNA-ZNF638 sequence were screened using the SRAMP procedure (http://www.cuilab.cn/sramp, accessed on 11 July 2021).
Overexpression/siRNA interference and cell co-cultivation
The SHF-stem cells (stored in our laboratory) were used in the overexpression/siRNA interference analysis of m6A-circRNA-ZNF638. The overexpression analysis of circRNA-ZNF638 (or its mutant) was performed in SHF-stem cells of cashmere goats, where the mutations of circRNA-ZNF638 were generated using the QuikChange Lightning Multi Site-Directed Mutagenesis Kit (Agilent, Technologies, Santa Clara, CA, USA) following the manufacturer’s recommendations. Overexpression vectors were constructed as described protocol in previous publication [13]. In brief, the pcDNA3.1 (+) circRNA mini-vector (Addgene, Cambridge, MA, USA) was utilized in overexpression assay. The SHF stem cells were transiently transfected with the recombinant pcDNA3.1 (+) circRNA-ZNF638 (or its mutant), which was carried out by the Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA). For the knockdown assay of circRNA-ZNF638, we designed three specific siRNAs (si-circR-1, si-circR-3, and si-circR-3) that potentially targeted the back splice region of circRNA-ZNF638. The three siRNA sequences are si-circR-1: 5′-AT GTAGTCATTTGAACTTTGTG-3′, si-circR-2: 5′-GAATG TAGTCATTTGAACTTTG -3′, and si-circR-3: si-circR-3: 5-TCATTTGAACTTTGTGTTATTC-3′. These siRNAs were synthesized commercially by GenePharma Co., Ltd. (Shanghai, China). Using the siRNAs Lipofectamine RNAiMAX kits (Invitrogen, Shanghai, China), the siRNAs were transiently transfected into SHF stem cells according to the manufacturer’s recommendations.
The transfected SHF stem cells were co-cultured with DPCs (stored in our laboratory) for induced activation in a transwell device as described by Yan and colleagues [4]. In brief, the DPCs (passage 3) of cashmere goats were seeded on a transwell insert, which was added to a six-well plate seeded with SHF stem cells (passage 3) of cashmere goats. Subsequently, in fresh DMEM/F12 medium (Hyclone, Logan, UT, USA; supplemented with 10% fetal bovine serum), the cells were non-contactingly co-cultured under a humidified atmosphere at 37°C with 5% CO2. The culture media was replaced every two days. After post-transfection 48 h, the transfected cells were collected, and the transfection efficiency was evaluated by RT-qPCR assay. The transfection efficiency was above 70%.
Primer designing and RT-qPCR assay
For m6A-circRNA-ZNF638, divergent primer pair was cited from the previous study [2], and for the methylation immunoprecipitation (Me-RIP) with real-time quantitative polymerase chain reaction (RT-qPCR) of m6A-circRNA-ZNF638 and mRNAs of the related genes, convergent primer pairs were designed using the Primer Premier 5.0 program (http://www.premierbiosoft.com). Whereas, for miRNAs, the sense primer of each miRNA was designed based on its mature sequence that was retrieved from the miRNA database: miRNAsong (https://www2.med.muni.cz/histology/miRNAsong/index.php), and the anti-sense primers are universal to all miRNAs that were obtained from the kits (TaKaRa, China). The primer pair of U6 was cited from the previous study [20]. The details of all primers are presented in Table 1.
For the expression analysis of m6A-circRNA-ZNF638 (or it’s Me-RIP-qPCR) and mRNAs of implicated genes, the M-MuLV cDNA synthesis kit (Sangon, Shanghai, China) was used for synthesizing the first strand cDNA with random primers. Whereas for miRNA expression analysis, the One-Step PrimeScript microRNA cDNA synthesis kit (TaKaRa, China) was used for synthesizing the first strand cDNAs. The RT-qPCR assay carried out in a light Cycler 480 real-time PCR system (Roche Diagnostics, Germany). In a 25 μL final volume, RT-qPCR amplication was performed using TB Green Premix Ex Taq II (Tli RNaseH Plus; TaKaRa, Chnia) containing 1.0 μL each primer (10 μM), 2.0 μL first-strand cDNA solution, 12.5 μL TB Green Premix Ex Taq II (Tli RNaseH Plus; TaKaRa, Chnia), and 8.5 μL PCR-grade ddH2O water. We used the following thermal cycling parameters: a single cycle (95°C for 30 s) followed by 40 cycles: 95°C for 5 s, 53°C to 60°C (Table 1) for 30 s, and 72°C for 30 s. The m6A relative enrichment of m6A-circRNA-ZNF638 was normalized to the input, and the mRNA relative expression of the implicated genes was calculated with the 2−ΔΔCt method.
Methylation Immunoprecipitation of m6A-circRNA-ZNF638 (Me-RIP) in SHF stem cells
The Me-RIP analysis of m6A-circRNA-ZNF638 was conducted as described protocol by Chen and colleagues [21]. Briefly, 100 μg total RNA was treated with RNase R (Geneseed, Guangzhou, China), and further concentrated using Monarch RNA Cleanup Kit (NEB, Ipswich, MA, USA). Subsequently, RNA was fragmented at 94°C for 3 min using the NEBNext Magnesium RNA Fragmentation Module (NEB, USA), and further concentrated with the Monarch RNA Cleanup Kit (NEB, USA). The fragmented product of 2 μg was reserved as input control. Half fragmented RNA was incubated anti-m6A antibody of 2 μg (Synaptic Systems, Gottingen, Germany) or IgG of 2 μg (Cell Signaling Technology, Danvers, MA, USA) at 4°C for 4 h. After pre-washing, the Dynabeads Protein A (Thermo Scientific, Rockford, IL, USA) was subjected to incubation with the complex of RNA and antibody at 4°C for 2 h. The RNA was extracted with the RNAiso kit (TaKaRa, China), and the expression of m6A-circRNA-ZNF638 of each sample was analyzed via the RT-qPCR assay.
Dual-luciferase reporter assays
The dual-luciferase reporter assay was carried out as described in previous publication [22]. In brief, Luciferase reporters were generated by cloning circRNA-ZNF638 (circRNA-ZNF638-WT) or its mutant (circRNA-ZNF638-MUT) into pGL3-basic vectors (Promega, Madison, WI, USA). The mutant of circRNA-ZNF638 (circRNA-ZNF638-MUT), that was mutated within the potential binding sites of miR-361-5p seed region, were generated using the QuikChange Lightning Multi Site-Directed Mutagenesis Kit (Agilent, Technologies, USA) following the manufacturer’s recommendations. Also, the 3′-untranslated region (3′-UTR) fragment of cashmere goat Wnt5a mRNA was amplified, which contained the potential binding site of miR-361-5p and was ligated to the pGL3 basic vector (Promega, USA). And then, the SHF-stem cells (passage 3) of cashmere goats were prepared and transfected with the generated reporter vectors via the use of Lipofectamine 2000 (Invitrogen, USA). After post-transfection 48 h, the transfected cells were collected, and the transfection efficiency was evaluated by RT-qPCR assay. The transfection efficiency is above 70%. The transfected cells were cultured under the above-mentioned conditions. After 48 h, the cell lysates were harvested and luciferase activities were examined using the Dual-Luciferase Reporter Assay System (Promega, USA). The ratio of firefly to renilla luciferase activity was measured to eliminate possible bias in transfection efficiency.
Data statistical analysis
Statistical analysis of obtained data was performed with the SPSS 17.0 program (SPSS Inc., Chicago, IL, USA). The data was presented as mean±standard deviation. The comparison for potential difference between two groups was performed with the Student’s t-test. The resulting p-values less than 0.05 were considered statistically significant.
RESULTS AND DISCUSSION
Molecular characterization of m6A-circRNA-ZNF638 in SHFs of cashmere goats
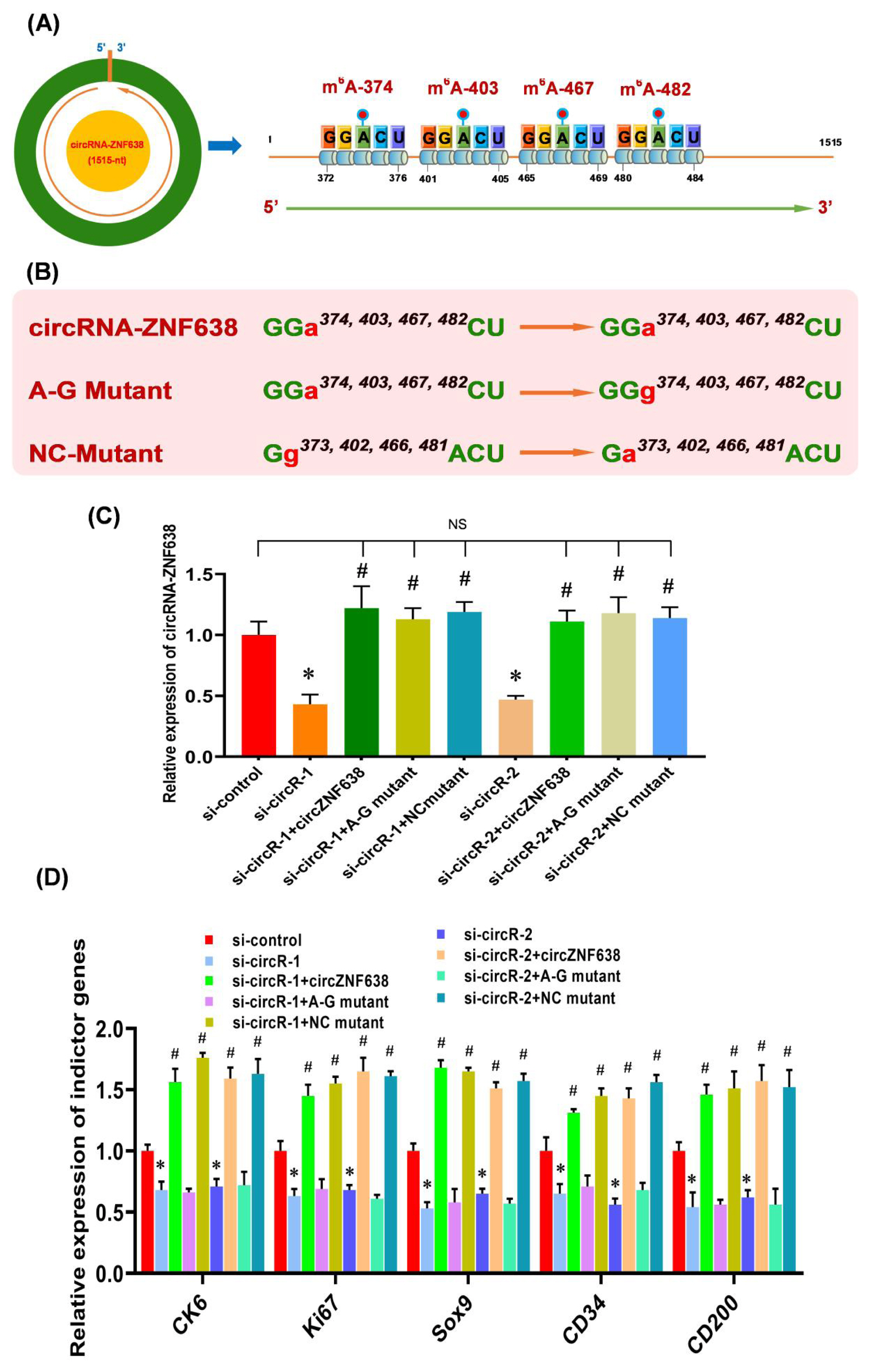
Here, the analyzed m6A-circRNA-ZNF638 has been identified from cashmere goat SHFs in our previous study [2]. Through an alignment of its linearized sequence into the goat genome (https://www.ncbi.nlm.nih.gov/genome/?term=goat,assembly,ARS1), it was revealed that the m6A-circRNA-ZNF638 was transcribed from the antisense strand of goat ZNF638 gene (Figure 1A). In fact, the goat ZNF638 gene contains multiple exons including exons 1 to 28, which was well annotated in NCBI database (https://www.ncbi.nlm.nih.gov). In detail, the m6A-circRNA-ZNF638 is formed via a reverse splicing of the antisense strand of entire exon 2 of the goat ZNF638 gene with position nos. 13101422–12102936 of the NC_030818.1 sequence on chromosome 11 (Figure 1A), and it is 1515-nt in spliced length.
Through in-silicon analysis, we also noted that m6A-circRNA-ZNF638 contained potential target sites of multiple miRNAs, including miR-103-3p, miR-107-3p, miR-361-5p, miR-140-3p, and miR-194 (Figure 1B), which implies that m6A-circRNA-ZNF638 in SHFs of cashmere goats may exert its biological functions through miRNA-mediated pathways as reported in recent publications [13–15]. On the other hand, the four m6A sites within m6A-circRNA-ZNF638 sequence were revealed to have the same motif of GGACU, which is consistent with the m6A motif: RRACH (R, A/G; H, A/C/U) reported in linear RNA molecules (Figure 1B) [23].
Expression characterization of m6A-circRNA-ZNF638 in SHF bulge of cashmere goats and its effects on the induced activation of SHF-stem cells
For revealing the expression characterization of m6A-circRNA-ZNF638 in the SHF bulge of cashmere goats, two main phases of SHF cycle were investigated including anagen and telogen [1]. As observed from Figure 2A, m6A-circRNA-ZNF638 exhibited significantly higher expression in the anagen SHF bulge compared with that of telogen. As well known, at anagen bulge of cashmere goats, the SHF-stem cells are under vigorous status of induced activation from the DPC signal stimulation to drive the regeneration and growth of SHFs [3]. As observed from Figure 2B, this was also supported well by the tested results from this study, where several indicator genes upon induced activation of SHF-stem cells exhibited significantly higher expression at anagen bulge compared with those at telogen including keratins CK6, Ki67, Sox9, CD34, and CD200 [5,24,25]. Thus, we speculate that m6A-circRNA-ZNF638 may be significantly implicated in the induced activation event of SHF-stem cells in cashmere goats.
To confirm this hypothesis, we carried out the knockdown experiment of m6A-circRNA-ZNF638 in SHF-stem cells of cashmere goats through introducing siRNA inference molecules. Three independent siRNAs were designed including si-circZNF638-1, si-circ ZNF638-2, and si-circ ZNF638-3, and their knockdown efficiencies were verified in SHF-stem cells of cashmere goats. As shown in Figure 2C, the si-circZNF638-1 and si-circ ZNF638-2 was more efficient in knockdown of m6A-circRNA-ZNF638 compared with that of si-circ ZNF638-3 (Figure 2C). Therefore, the si-circZNF638-1 and si-circ ZNF638-2 were selected and utilized in further experiments. As shown in Figure 2D, the si-circZNF638-1/-2-mediated knockdown of m6A-circRNA-ZNF638 resulted in a significant decrease in the expression of the analyzed indicator genes upon induced activation of SHF-stem cells in comparison to those of the control group (si-control) (Figure 2D). Thus, it can be inferred that m6A-circRNA-ZNF638 may be essentially implicated in contributing the induced activation of SHF-stem cells in cashmere goats through certain unknown mechanisms.
Recently, several studies showed that circRNAs were essentially implicated in the differentiation of SHF-stem cells into hair follicle lineage in cashmere goats through loss-of-function experiments [13–15], but little information is available about the functional role of m6A modified circRNAs in SHF-stem cells of cashmere goats. Also, it is unclear how circRNAs epigenetically modulate the induced activation and cell-fate decisions of SHF-stem cells of cashmere goats. Interestingly, here, we verified the m6A modification of circRNA-ZNF638 in SHF-stem cells of cashmere goats by performing a methylated RNA immunoprecipitation (Me-RIP) assay (Figure 2E), which suggests that m6A-circRNA-ZNF638 may play functional roles in contributing to induced activation of SHF-stem cells through m6A modification-dependent pattern.
The m6A modification of circRNA-ZNF638 is implicated in its contributing to induced activation of SHF-stem cells
Recent studies have suggested that the m6A modifications are heavily implicated in the functional exertion of m6A-circRNAs [11,16]. This promotes us ask whether the observed functional role of m6A-circRNA-ZNF638 in contributing the induced activation of SHF-stem cells of cashmere goats was achieved through m6A-dependent pattern. As described in our previous publication [2], four m6A sites were revealed within circRNA-ZNF638 sequence, and they were named as m6A-374, m6A −403, m6A −467, and m6A −482, respectively (Figure 3A). Therefore, we constructed the expression vectors of circRNA-ZNF638 (wild type, WT) and its mutant expression vectors (mutant type, MUT), in which the adenine residues embedded within the m6A motifs (GGACU) were replaced with guanine residues (A-G mutation) (Figure 3B). We also constructed the mutants of circRNA-ZNF638 in which the four guanine residues (G373, G402, G466, and G481) within the m6A motifs (GGACU) were replaced with unmethylated adenine residues (G-A mutation) as control mutant (NC mutant) (Figure 3B) as reported by Yang and colleagues [26].
To evaluate the relationship between m6A modification and the functional role of circRNA-ZNF638 in SHF-stem cells, we overexpressed circRNA-ZNF638 (WT), the m6A-deficient A-G mutant (MUT) or the m6A-decorated mutant (NC mutant) with equal amounts in circRNA-ZNF638-knockdown SHF-stem cells and used them in vitro induced activation assays. We found that the mutations were not implicated in expression levels of circRNA-ZNF638, as there was no significant difference in circRNA-ZNF638 expression between the wild type- and mutant-transfected SHF-cells (Figure 3C). On the other hand, we also noted that the expression level of circRNA-ZNF638 was restored in circRNA-ZNF638-knockdown SHF-stem cells through the introduction of either wild type- circRNA-ZNF638 or its mutants (Figure 3C).
Next, we analyzed the expression changes of the indictor genes upon induced activation of SHF-stem cells in the transfected cells. As observed from Figure 3D, the ‘si-circR-1/2+circRNA-ZNF638’ cell lines, not the ‘si-circR-1/2+A-G mutant’ cells, exhibited a significantly higher expression of the analyzed indictor genes, compared with the si-circR-1/2 cell lines (Figure 3D). Notably, there is no significant difference in circRNA-ZNF638 expression levels between ‘si-circR-1/2+circRNA-ZNF638’ and ‘si-circR-1/2+A-G mutant’ cells (Figure 3C). Thus, we can rule out the possibility that the observed expression changes of the analyzed indictor genes between two cells types (si-circR-1/2+circRNA-ZNF638’ and ‘si-circR-1/2+A-G mutant’) were related with the circRNA-ZNF638 expression abundance. On the other hand, as expected, the NC mutant (m6A-decorated mutant) restored the indictor expression level of circRNA-ZNF638-deficient cells (Figure 3D). Taken together, these results suggest that the m6A modifications are necessary for circRNA-ZNF638 in contributing to the induced activation of SHF-stem cells in cashmere goats. A highly similar functional mechanism was also reported in mouse embryonic stem cells (mESCs), where the internal m6A modification was verified to positively participate in the functional role of linc1281 in the differentiation potential of mESCs [26].
M6A-circRNA-ZNF638 functions as miR-361-5p sponge and may negatively regulate its expression
Previous studies have demonstrated that circRNAs can function as natural miRNA sponges to control gene expression through posttranscriptional regulation patterns [27]. Therefore, we screened the candidate target miRNAs of m6A-circRNA-ZNF638 via in-silico analysis. As shown in Figure 4A, five candidate miRNAs were found to have the potential binding sites within m6A-circRNA-ZNF638 sequence, including miR-103-3p, miR-107-3p, miR-361-5p, miR-140-3p, and miR-194 (Figure 4A). To determine which miRNAs were sequestered by m6A-circRNA-ZNF638, a luciferase reporter was constructed which contains full-length circRNA-ZNF638 in the 3′-UTR of luciferase gene. The reporters were co-transfected into SHF-stem cells with the single candidate miRNA mimics, respectively. As a result, in comparison to the control mimics, only the miR-361-5p led to a significantly decrease in the luciferase activities of the m6A-circRNA-ZNF638 reporters, but not for miR-103-3p, miR-107-3p, miR-140-3p, and miR-194 (Figure 4B). To further define the specifically binding of miR-361-5p to the predicted target site within m6A-circRNA-ZNF638, a mutant luciferase report (circRNA-ZNF-MUT) was constructed which harbors an antisense mismatch of 7-nt within seed target site of miR-361-5p (Figure 4C). When co-transfected with miR-361-5p, we found that the mutant reports were counteractive to the miR-361-5p-driven reporter suppression (Figure 4D). Thus, these findings suggest that m6A-circRNA-ZNF638 serves as a sponge via direct binding with miR-361-5p in SHF-stem cells of cashmere goats.
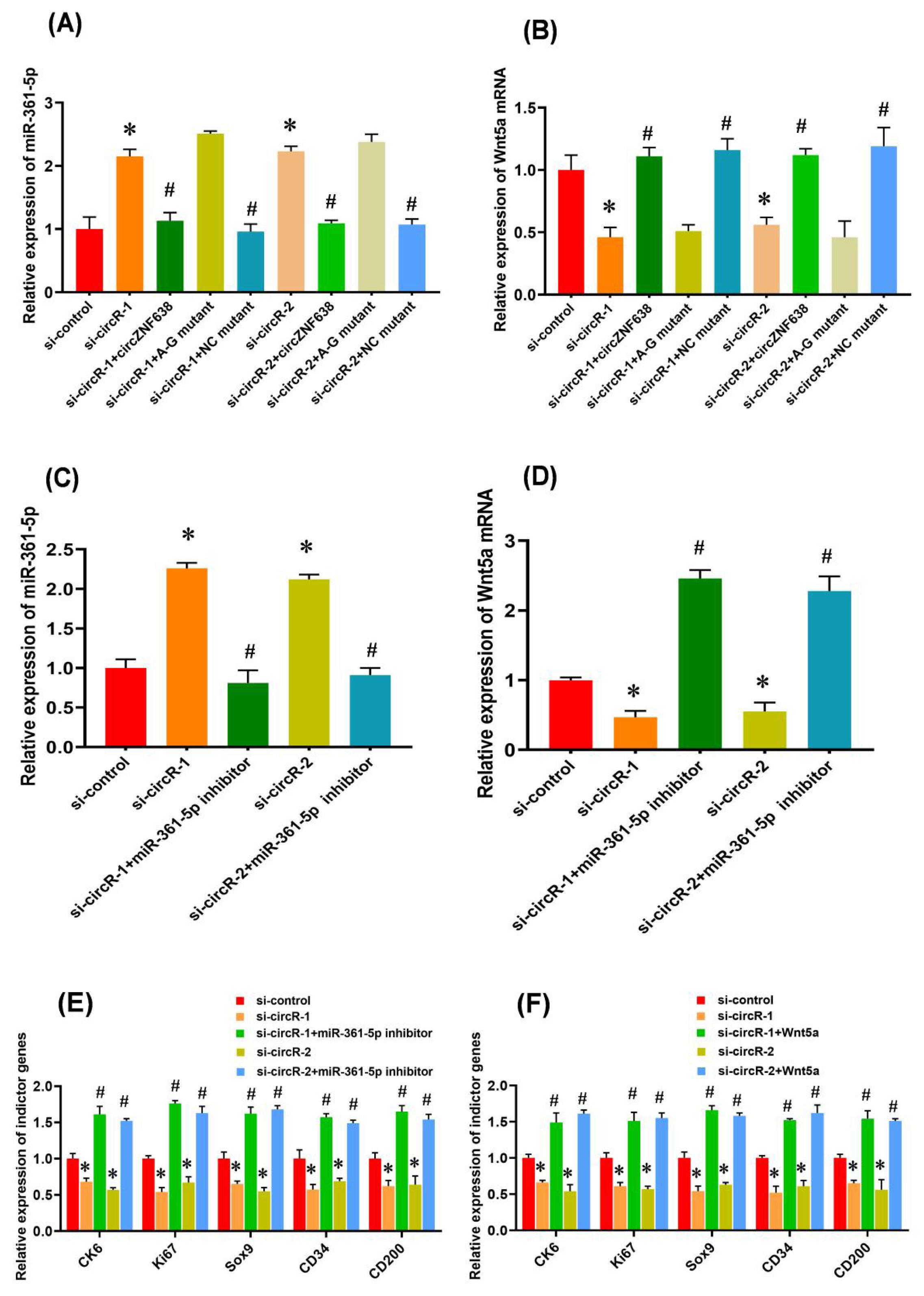
In addition, we found that the knockdown of m6A-circRNA-ZNF638 resulted in a significant increase of miR-361-5p expression in SHF-SCs (Figure 4E). However, neither overexpression nor knockdown of miR-361-5p had a significant effect on the expression of m6A-circRNA-ZNF638 in SHF-SCs of cashmere goats (data not shown). Therefore, we speculate that m6A-circRNA-ZNF638 may regulate the expression of miR-361-5p in SHF-stem cells. This regulatory mode is highly similar to that reported in investigation on the differentiation of cashmere goat SHF-stem cells into hair follicle lineage, where circRNA-0100 was revealed to directly bind with miR-153-3p, and negatively regulated its expression [15]. Also, in research on invasion and metastasis of colorectal cancer, Han and colleagues found that circLONP2 bound with miR-17, and negatively regulated its expression [28].
M6A-circRNA-ZNF638 up-regulates the expression of Wnt5a in SHF-stem cells of cashmere goats through miR-361-5p mediated pathway
In previous studies, it was demonstrated that the Wnt signaling pathway was heavily implicated in the initiation of hair follicle development [29]. The Wnt5a, a member of Wnt family, has been shown to be critical for controlling the fate of hair follicle cells [29]. Moreover, it is believed that Wnt5a can promote the activation of human hair follicle stem cells leading to the onset of anagen stages [30]. These findings drove us to ask whether the Wnt5a may be implicated in the observed effect of m6A-circRNA-ZNF638 on the induced activation of SHF-stem cells through miR-361-5p mediated pathway. Therefore, we further investigated the expression change of Wnt5a in m6A-circRNA-ZNF638 knockdown SHF-stem cells. As shown in Figure 4F, after the knockdown of m6A-circRNA-ZNF638, the relative expression level of Wnt5a was significantly down-regulated in SHF-SCs of cashmere goats (Figure 4F). These results suggested that m6A-circRNA-ZNF638 may be positively related to the Wnt5a expression in the SHF-stem cells.
It was widely accepted that circRNA can act as miRNA sponge to reduce the active miRNAs, which disinhibits the expression of the miRNA target genes at the post-transcriptional level [31]. Here, we confirmed that m6A-circRNA-ZNF638 sequestered miR-361-5p (Figure 4B, 4C, and 4D), and positively regulated the expression of Wnt5a in SHF-stem cells (Figure 4F). These results raised a most likely mechanism that m6A-circRNA-ZNF638 may positively regulate the Wnt5a expression in SHF-stem cells via miR-361-5p mediated pathway. Thus, we performed an in-silico screen for potential binding sites of miR-361-5p within the 3′-UTR region of Wnt5a mRNA sequence. As shown in Figure 4G, a potential binding site of miR-361-5p was harbored within the 3′-UTR region of Wnt5a mRNA with a 7 mer binding type of seed region (Figure 4G). To confirm this interaction between miR-361-5p and the 3′-UTR region of Wnt5a mRNA, we constructed a luciferase reporter of 3′-UTR region of Wnt5a mRNA containing the potential binding site for miR-361-5p, and co-transfected the constructed reporter into the SHF-stem cells with si-RNAs of m6A-circRNA-ZNF638. As shown in Figure 4H, the si-circR-1/2 mediated knockdown of m6A-circRNA-ZNF638 (si-circR-1/2) in SHF stem cells significantly decreased the relative luciferase activity of Wnt5a mRNA 3′-UTR compared with siRNA control (si-control) (Figure 4H). In further order to verify the specifically binding of miR-361-5p to the predicted target site within the 3′-UTR region of Wnt5a mRNA, a mutant luciferase report (Wnt5a mRNA 3′-UTR-MUT) was constructed which harbors an antisense mismatch of 7-nt within seed target site of miR-361-5p (Figure 4I). When co-transfected into SHF-stem cells with miR-361-5p, we found that relative luciferase activity of mutant reports (Wnt5a mRNA 3′-UTR-MUT) had no significant change compared with that of miRNA control minics (Figure 4J). These results indicated the specifically target binding of miR-361-5p to 3′-UTR region of Wnt5a mRNA.
In fact, functionally, it was demonstrated that circRNAs could play roles in cells through multiple pathways and mechanisms, such as, sponging miRNAs [27], regulating transcription of host gene [32–35], controlling RNA transport [36], and serving as template for protein translation [11]. Here, we showed that m6A-circRNA-ZNF638 sponged miR-361-5p to upregulate the expression of Wnt5a in SHF-stem cells of cashmere goats. However, the other functional roles of m6A-circRNA-ZNF638 in SHF-stem cells of cashmere goats should be further investigated, such as its potential function of regulating ZNF638 transcription, and encoding a protein which may have essentially significant to the physiological function of SHF-stem cells in cashmere goats.
The m6A modification is necessary for the circRNA-ZNF638-mediated regulatory effects through miR-361-5p/Wnt5a axis
There is evidence that m6A modifications within RNA molecules are significantly implicated in various cellular biological processes, such as miRNA biogenesis initiation [35], pre-mRNA splicing [36], and circRNA translation initiation [11]. Although, the functional significance of m6A modification within circRNAs still needs to be clarified, an m6A -dependent model was reported on the interaction between lncRNA and miRNA in which m6A modification of linc1281 was found to be required for its directly binding with let-7 family miRNAs [26]. This promotes us ask whether m6A modification of circRNA-ZNF638 is necessary for its regulatory effect on the induced activation of SHF stem cells through miR-361-5p/Wnt5a axis. To define this hypothesis, we first tested the expression level of miR-361-5p in ‘siRNA-circR-1/2+circRNA-ZNF638’, ‘siRNA-circR-1/2+ circRNA-A-G mutant’, and ‘siRNA-circR-1/2+NC mutant’ SHF-stem cells. We noted that ‘siRNA-circR-1/2+circRNA-ZNF638’, and ‘siRNA-circR-1/2+NC mutant’ cells exhibited decreased miR-361-5p expression compared with ‘si-circR-1/2’ cells (Figure 5A). However, the ‘siRNA-circR-1/2+A-G mutant’ cells still exhibited high miR-361-5p expression levels (Figure 5A), despite there is no significant difference in circRNA-ZNF638 transcript levels among these different treated cell types (Figure 3C). We further examined the expression level of Wnt5a mRNA in ‘siRNA-circR-1/2+ circRNA-ZNF638’, ‘siRNA-circR-1/2+circRNA-A-G mutant’, and ‘siRNA-circR-1/2+NC mutant’ SHF-stem cells. We found that the ‘siRNA-circR-1/2+A-G mutant’ cells exhibited decreased Wnt5a mRNA expression compared with ‘si-circR-1/2’ cells (Figure 5B). However, the ‘siRNA-circR-1/2+circRNA-ZNF638’, and ‘siRNA-circR-1/2+NC mutant’ cells still displayed high Wnt5a mRNA levels (Figure 5B). Taken together, it appears to become apparent that the m6A modification is necessary for the circRNA-ZNF638-mediated regulatory effects through miR-361-5p/Wnt5a axis.
In fact, it is widely accepted that the binding between circRNAs (or lncRNAs) and miRNAs is usually driven through a sequence-based complementary base pairing model in which whether regulators mediate this binding is still unclear [14,26]. Here, we showed that m6A positively mediated the binding between m6A-circRNA-ZNF638 and miR-361-5p, which is revealed by m6A-deficient A-G mutant of circRNA-ZNF638 that abolished the binding of circRNA-ZNF638 with miR-361-5p (Figure 5A and 5B). On the other hand, previous investigations on m6A peak of mRNA and miRNA binding sites have suggested a possible mechanism via which m6A might cooperate or compete with miRNAs to ultimately regulate the expression of target mRNAs [37]. In this study, we suggested a model that circRNAs harbor both m6A peaks and miRNA binding sites in which m6A modification cooperate with miRNA to drive the circRNA-mediated ceRNA regulation, thereby contributing to the induced activation of SHF-stem cells of cashmere goats. Although it remains unclear that whether m6A modification alters the local structure of circRNA-ZNF638 as reported in a previous publication [38] thereby driving the binding with miR-361-5p, our results provide significant insights into the m6A mediated regulatory model of circRNA in SHF-stem cells of cashmere goats.
The miR-361-5p/Wnt5a axis restores the induced activation of SHF stem cells with m6A-circRNA-ZNF638-deficient
And then, we asked whether the miR-361-5p/Wnt5a axis is responsible for the m6A-circRNA-ZNF638-mediated regulation on the induced activation of SHF-stem cells. For this purpose, miR-361-5p inhibitor was transfected into m6A-circRNA-ZNF638 knockdown cells. As expected, the miR-361-5p expression was significantly decreased by its inhibitor in circRNA-ZNF638 knockdown SHF-stem cells (Figure 5C). Coupled with the observation, Wnt5a mRNA level was significantly increased by the miR-361-5p inhibitor in circRNA-ZNF638 knockdown SHF-stem cells (Figure 5D). Interestingly, we found that concomitant inhibition of miR-361-5p was sufficient to rescue the impaired inducted activation of SHF-stem cells after m6A-circRNA-ZNF638 knockdown, which can be determined by the significant up-regulation of the indicator genes on induced activation of SHF-stem cells (Figure 5E).
On the other hand, since Wnt5a mRNA was revealed to bind directly with miR-361-5p (Figure 4G-4J), and showed decreased levels upon m6A-circRNA-ZNF638 knockdown (Figure 5B). We also examined whether Wnt5a contributed to the underlying mechanism of the m6A-circRNA-ZNF638/miR-361-5p model. We overexpressed Wnt5a in SHF-stem cells with m6A-circRNA-ZNF638 knockdown. As shown in Figure 5F, the overexpression of Wnt5a restored the inducted activation of SHF-stem cells caused by m6A-circRNA-ZNF638 deficiency, which can be determined by the significant up-regulation of the indicator genes on induced activation of SHF-stem cells (Figure 5F).
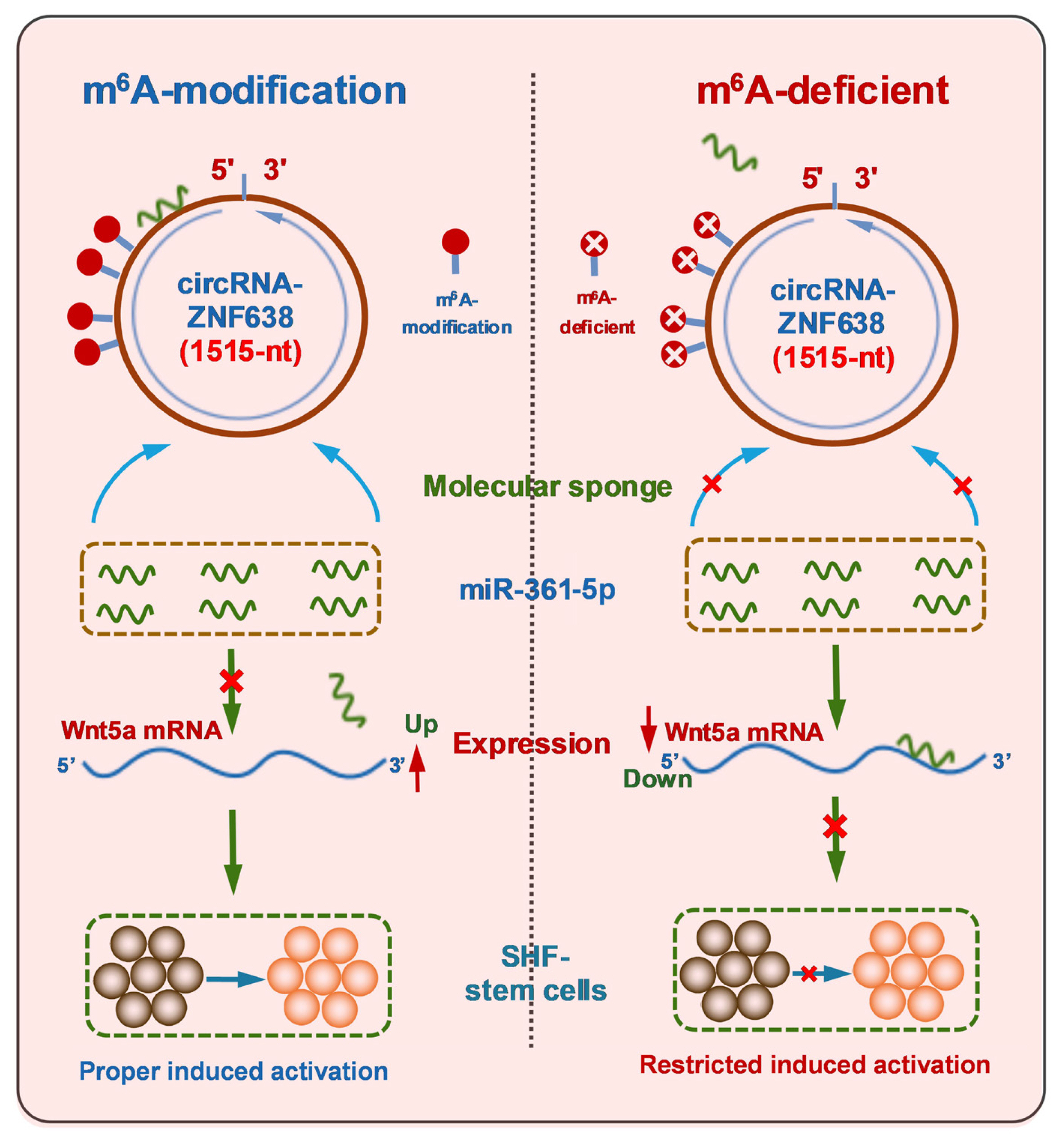
Previously, although it was thought that Wnt5a, as a member of non-canonical Wnt family, could inhibit the canonical Wnt/β-catenin signaling that was essentially involved in the initiation of hair follicle development [29], and it is not yet known whether non-canonical Wnt5a is directly implicated in the induced activation event of SHF-stem cells of cashmere goats. However, it was reported that Wnt5a also could activate canonical Wnt/β-catenin signaling during mouse embryonic development [39]. Moreover, compared with catagen and the telogen, Wnt5a exhibited the highest levels at the anagen hair follicles with being prominently located in the matrix, precortex cells, inner root sheath, outer root sheath and the dermal papilla [29]. Also, it was found that overexpression of Wnt5a in DPCs led to a significant decrease in expression of the related genes involved in maintaining cell quiescent state [40]. In addition, Wnt5a can serve as both SHH signaling target and Notch/CSL signaling mediator in the morphogenesis and differentiation of hair follicles, respectively [41]. Here, we found that m6A modification of circRNA-ZNF638 of cashmere goats is required for the induced activation of SHF stem cells through miR-361-5p/Wnt5a axis (Figure 6). Nevertheless, it is worth noting that, here, we conducted the experiment in SHF-stem cells in vitro. Therefore, the revealed functional mechanism of m6A-circRNA-ZNF638 above should be further confirmed in SHF-stem cells in vivo.



 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







