





INTRODUCTION
Clostridium perfringens, gram-positive and spore-forming anaerobe, is the third common cause of foodborne illness in United States [1]. Also, this bacterium is responsible for the symptoms, such as abdominal cramps and severe diarrhea, caused by ingestion of food which is contaminated in large number of C. perfringens, especially proteinaceous foods, meat, and poultry [2]. Besides, C. perfringens generally inhabits in a human gut, and its prevalence in the gut increases, as humans get old [3]. A significant proportion of this bacterium in gut microbiota may be associated with autism in children and neurological disorders, such as neuromyelitis optica [4ŌĆō6]. The virulence of C. perfringens attributes to its toxins, especially C. perfringens enterotoxin (CPE) produced during its sporulation in intestines [7]. Due to its ability to bind to claudin receptors, which are junction proteins in intestines, it is known that CPE creates pore on the cell membrane and results in cell death, causing diarrhea and abdominal pain [7]. Thus, this bacterium needs to be controlled. However, there are no edible materials which are applicable to human for destroying C. perfringens specifically inhabits in intestines.
In previous studies, glycated milk proteins, such as whey protein glycated with galactose and glucose, were studied for anti-inflammatory activities and their protective effects against oxidative stress [8,9]. Furthermore, fermented Maillard reaction products (FMRPs), which were produced from glycated milk proteins, have recently been introduced for their physiological functions, such as antioxidant, antihypertensive, antithrombotic, and hepatoprotective properties [10,11]. It was reported that antimicrobial activities of Maillard reaction products vary according to the kinds of amino acids [12]. Moreover, Maillard reaction products, which were derived from the foods, such as coffee, beer, and wine, showed different antimicrobial activities [13,14]. Meanwhile, the antimicrobial activity of FMRPs has never been investigated yet to date. Moreover, the effect of Maillard reaction products against the bacteria inhabiting gastrointestinal tract was not elucidated. In order to reduce incidence of diseases due to the pathogenicity of intestinal microbes, and risks such as antibiotics and stress tolerance, it is necessary to study FMRPs possessing physiological and antimicrobial activities. Therefore, the objective of this study was to evaluate the antimicrobial activity of FMRPs against C. perfringens and to elucidate the mode of FMRPs for antimicrobial activity.
MATERIALS AND METHODS
Preparation of fermented Maillard reaction products
FMRPs were prepared with whey protein (Davisco Foods International Inc., Le Sueur, MN, USA), sugars (galactose or glucose), and lactic acid bacteria (Lactobacillus rhamnosus 4B15 or Lactobacillus gasseri 4M13) (Table 1). For Maillard reaction, 50 mg/g of whey protein isolate and 25 mg/g of sugar (galactose or glucose) were mixed in deionized water, followed by shaking at 60 rpm in a pilot-scale instrument controlled by a pilot-scale pasteurization unit (Powerpoint International, Tokyo, Japan) at 65┬░C for 24 h. These Maillard reacted galactose and glucose were then fermented by L. rhamnosus 4B15 or L. gasseri 4M13 at 6 Log colony-forming unit (CFU)/mL at 37┬░C for 48 h to prepare FMRPs (Gal-4B15, Gal-4M13, Glu-4B15, and Glu-4M13). The cultures were then centrifuged at 2,000├Śg for 30 min (Hitachi CR-21G rotor R20A, Hitachi Ltd., Tokyo, Japan). Subsequently, the supernatants were transferred into aluminum containers, followed by freeze-drying using a lyophilizer (OPERON FDUT-8608, Gimpo, Gyeonggi, Korea) for 72 h. The freeze-dried FMRPs were powdered and stored at 4┬░C until further use.
Bacterial preparation
To evaluate antimicrobial effects of FMRPs on C. perfringens, 1 mL of C. perfringens strains NCCP15911, NCCP15912, NCCP10846, NCCP10858, NCCP10976, and NCCP10347 were inoculated in 10 mL cooked meat medium (Oxoid, Basingstoke, Hampshire, UK), and the cultures were placed in airtight containers with Anaerogen (Oxoid, UK), followed by incubation at 37┬░C for 24 h. Then, 0.1 mL aliquots of the cultures were inoculated into 9 mL fresh brain-heart infusion (BHI; BD Difco, Sparks, MD, USA), followed by anaerobic incubation at 37┬░C for 24 h. The subcultures were mixed, and then centrifuged at 1,912├Śg and 4┬░C for 15 min. The cell pellets were washed twice with phosphate buffered saline (PBS, pH 7.4; 0.2 g of KH2PO4, 1.5 g of Na2HPO4, 8.0 g of NaCl, and 0.2 g of KCl in 1 L of distilled water). The cell suspensions of C. perfringens were then diluted to OD600 = 0.01 with PBS to obtain 5 to 6 Log CFU/mL of inoculum.
Antimicrobial effect of fermented Maillard reaction products
To evaluate the antimicrobial effects of FMRPs on C. perfringens, a hundred microliters of C. perfringens inoculum and 900 ╬╝L of FMRPs (100 mg/mL; Gal-4B15, Gal-4M13, Glc-4B15, and Glc-4M13) were mixed in a well of a 24-well plate (SPL Life Sciences, Gyeonggi, Korea), and the plate was placed in an airtight container, containing Anaerogen (Oxoid, UK), followed by anaerobic incubation at 37┬░C for 24 h. The samples were then serially diluted with buffered peptone water (BPW; BD Difco, USA), and 100 ╬╝L of the diluents were spread-plated on tryptose sulfite cycloserine agar (TSC; Oxoid, UK) to enumerate C. perfringens cells.
Iron chelation activity
To evaluate the iron chelation activity by FMRPs, 100 ╬╝L of FMRPs (20 mg/mL), 600 ╬╝L of distilled water, and 100 ╬╝L of 0.2 mM iron (II) chloride tetrahydrate (FeCl2┬Ę4H2O, SAMCHUN chemical Co. Ltd., Seoul, Korea) were mixed, and the mixture reacted for 30 sec at room temperature. Subsequently, 200 ╬╝L of 0.1 mM Ferrozine (3-(2-Pyridyl)-5,6-diphenyl-1,2,4-triazine-p,pŌĆÖ-disulfonic acid monosodium salt hydrate; Sigma Aldrich, St. Louis, MO, USA) were suspended in the mixture, and the samples reacted for 10 min at room temperature. Thereafter, turbidity of the samples was measured at 562 nm with a UV-spectrophotometer (OPTIZEN 2120UV, Mecasys Co., Ltd., Daejeon, Korea), and iron chelation activity was calculated as follows;
Absorbance of blank solution was measured with distilled water instead of with samples [15].
Propidium iodide uptake assay
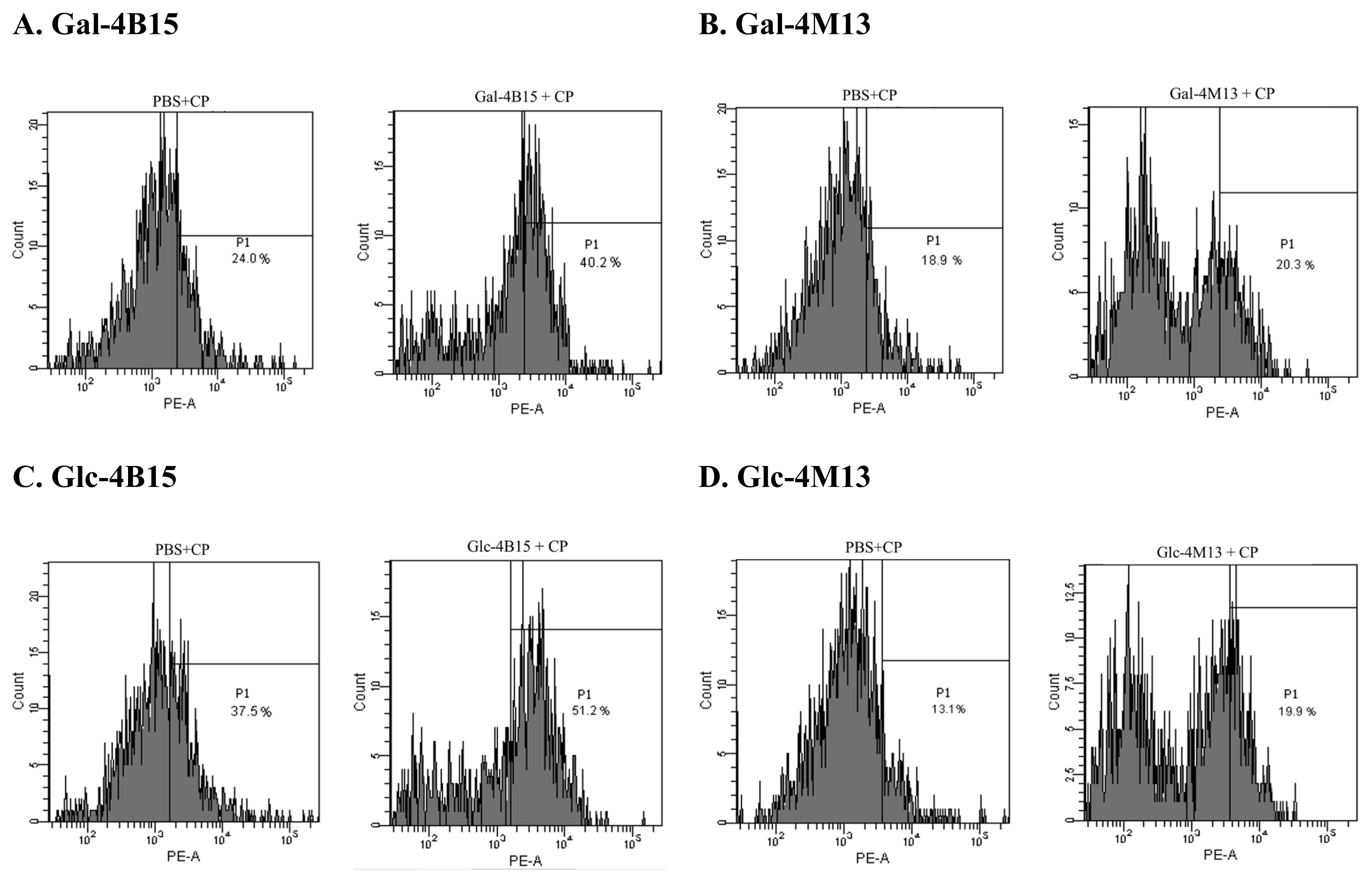
To observe the effects of FMRPs on bacterial cell permeability in short time, propidium iodide (PI) uptake assay for FMRPs-exposed C. perfringens was conducted according to the method described by Kim et al [16]. C. perfringens were cultured in cooked meat medium at 37┬░C for 24 h. The bacterial cultures were centrifuged at 10,000├Śg for 1 min at room temperature, and supernatants were discarded. One-milliliter portion of each FMRP sample (200 mg/mL) was transferred to the cell pellets and mixed, followed by incubation at 37┬░C for 60 min. Subsequently, 50 ╬╝L of C. perfringens incubated with FMRP were diluted with 950 ╬╝L of PBS in a FACS tube (BD Bioscience, San Jose, CA, USA), and 10 ╬╝L of PI solution (1 mg/mL) (Sigma Aldrich, USA) was then added to the diluted cultures. The mixtures were analyzed with FACSCanto II (BD Bioscience, USA), and PI activities for C. perfringens exposed to FMRPs were calculated, using CellQuest Pro software (BD Bioscience, USA). The percentages of FMRPs-treated bacteria figured in each histogram are relatively calculated to PBS-treated bacteria at a same designated point.
Morphological observation
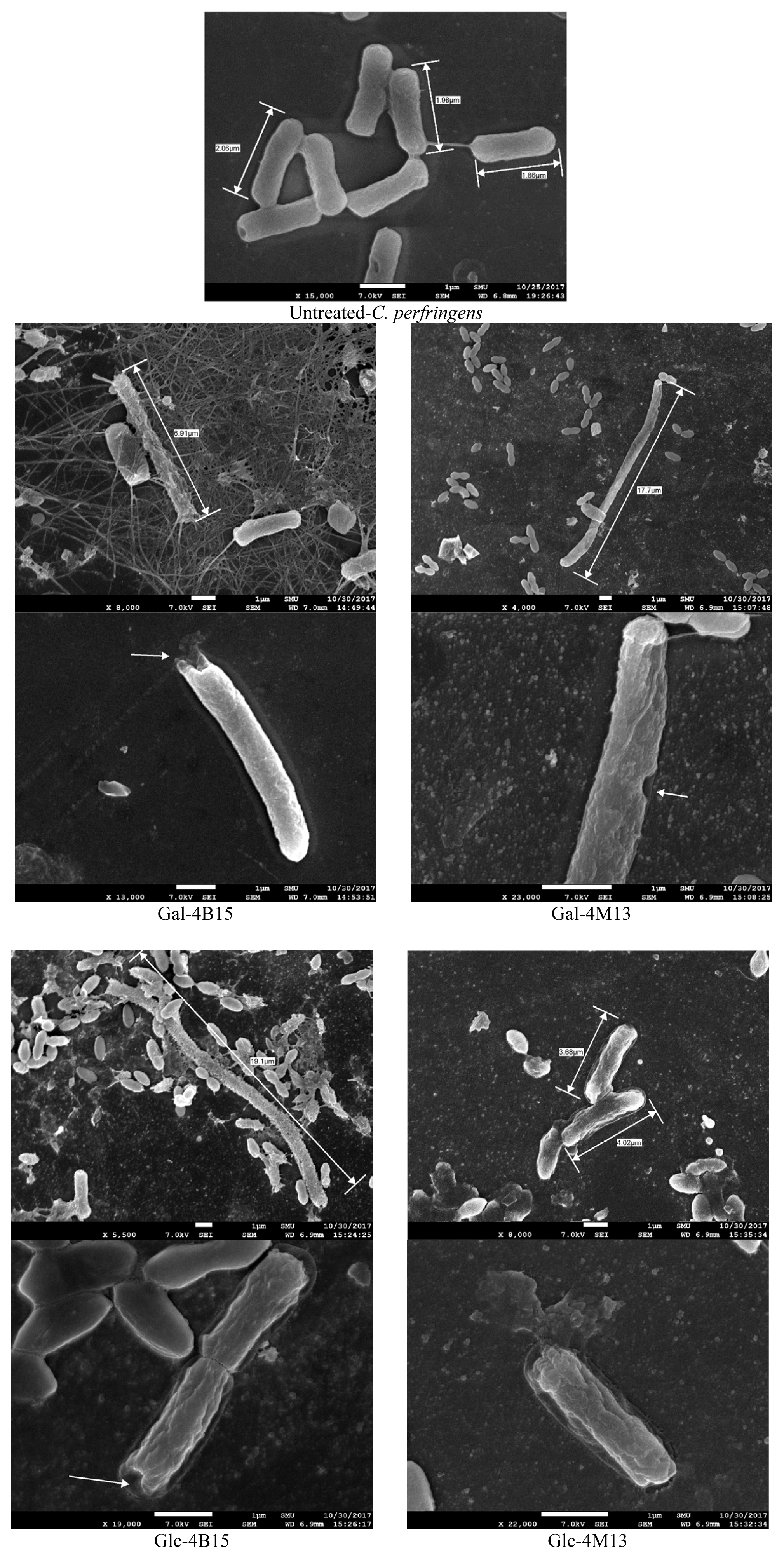
To elucidate the antimicrobial activity of FMRPs against C. perfringens, morphological observations were conducted, using field emission scanning electron microscope (FE-SEM; JEOL JSM-7600F, JEOL USA Inc., Peabody, MA, USA), according to the method described by Kim et al [16]. C. perfringens were cultured in 500 ╬╝L BHI containing the glass slides anaerobically at 37┬░C for 24 h with Anaerogen (Oxoid, UK). After incubation, the glass slides were removed from the cultures, and aseptically transferred to sterile 24-well flat-bottomed polystyrene plates (SPL, Korea). Thirty milliliters of 1.8% glutaraldehyde (Sigma-Aldrich Co., St. Louis, MO, USA) were carefully placed over the glass slides, and left for 30 min at room temperature for the reaction, followed by washing glass slides three times with 1 mL distilled water for 5 min. Subsequently, the glass slides were treated with 2% osmium tetroxide solution (Sigma Aldrich, USA) for 20 min at room temperature for a second fixation, followed by washing them three times with 1 mL distilled water for 5 min. The fixed bacterial cells on the glass slides were gradually dehydrated with 1 mL of 25%, 50%, 75%, 90%, and 100% ethyl alcohol for 5 min. Afterwards, the glass slides were thoroughly dried with 15 ╬╝L hexamethyldisilazane (Sigma-Aldrich, USA). The samples were eventually coated with a sputter coater (Cressington 108auto SEM sputter coater, Cressington Scientific Instruments Ltd., Watford, UK), and the bacterial cells were observed, using field emission scanning electron microscopy.
RESULTS AND DISCUSSION
Antimicrobial activities of fermented Maillard reaction products
C. perfringens cell counts in the FMRPs significantly decreased (p<0.05) at 37┬░C for 12 and 24 h (Figure 1), regardless of FMRPs used. Especially Gal-4B15 reduced C. perfringens cell counts below detection limit (10 CFU/mL) even after 12 h of incubation (Figure 1). This result indicates that FMRPs had significant antimicrobial effects against C. perfringens. The functional effects of FMRPs, such as antihypertension and prevention of cardiovascular disease, were found in other studies [10,11]. Taken together with the antimicrobial functions of FMRPs against C. perfringens, FMRPs may reduce intestinal C. perfringens and may be potential materials as health-promoting supplements. Although some fermented dairy products which show antimicrobial activity against C. perfringens, such as Kefir and probiotic yogurt, already exist, FMRPs are distinctive due to its antihypertensive and cardiovascular disease-preventive effects [17].
Iron chelation activity
Theoretically, high iron chelation activity indicates that iron as an essential nutrient for microorganisms in the environment is unavailable to bacteria, which in turn suppresses bacterial growth and decreases bacterial cell counts [18,19]. Gal-4M13 (23.87%) significantly showed high iron chelation activity compared to Glc-4M13 (0.50%) (p<0.05) (Table 2). This suggests that Gal-4M13 may have better iron chelation activities and thus, they may play an important role in inhibiting the growth by limiting uptake of iron in bacterial growth environments. However, only iron chelation activity is not affecting the antimicrobial effects of Gal-4M13 in this study, because Glc-4B15 and Glc-4M13 showed similar antimicrobial effects on C. perfringens, although they showed difference in iron chelation activity.
Propidium iodide uptake assay
In PI uptake assay, movement of peaks in a histogram, which represents the number of events detected at particular intensity, to the right indicates that the cell membrane is damaged. In this study, the movement of the peaks to right, and the change in the shape of the peaks and the number of the events were observed after the FMRPs treatment. Movement of fractions towards right X-axis indicates damages of bacterial cells via detection of wavelength of PI which binds to DNA leaked from damaged bacterial cells [20]. The events of treated C. perfringens increased from 24.0% to 40.2% by Gal-4B15 and from 18.9% to 20.3% by Gal-4M13. Glc-4B15 and Glc-4M13 treatments also increased the portions of C. perfringens from 37.5% to 51.2% and from 13.1% to 19.9%, respectively (Figure 2). This result indicates that FMRPs caused cell membrane damage, and it might be one of the modes to destroy C. perfringens cells. To date, physicochemical properties of dairy antimicrobial peptides, such as net charge, amphipathicity, hydrophobicity, etc., have shown effects on interaction with cell membranes and integration of the membranes with the peptides [21]. Therefore, it is assumed that peptides in FMRPs specifically interacted with the bacterial cell membranes and occurred the cell leakage.
Morphological analysis
The PI uptake assay showed that FMRP treatments caused cell membrane damages, which were confirmed visually by FE-SEM. FMRP-treated C. perfringens had an uneven cell membrane, and pores on the cell membrane surface (Figure 3). In addition, the lengths of the FMRP-treated bacteria increased from 2.0 ╬╝m to 12.0 ╬╝m in average and 19.1 ╬╝m in maximum for C. perfringens. Normally each bacteria species consistently maintains their classical bacterial shapes with their complex mechanisms genetically and biochemically [22]. Bacterial cells were found to elongate in order to adjust stressful environments [23]. Everis and Betts [23] also suggested that the length of Clostridium spp. increases when they are exposed to stress. When the bacterial cell is exposed to high solute and nutrition concentration, the osmotic forces change the mechanical integrity of bacterial cell and determine the shape [24]. Thus, the elongation of the bacterial cells in this study would be a result of the stressful environment by FMRPs. Taken together with the result of iron chelation activity and PI uptake assay, FMRPs would have antimicrobial activity as they occurred cell membrane damages and iron chelating properties simultaneously.
CONCLUSION
The FMRPs (Gal-4B15, Gal-4M13, Glc-4B15, and Glc-4M13), which are already known for the functional properties, showed significant antimicrobial activity against C. perfringens. The antimicrobial activity of FMRPs would be caused by iron chelation and bacterial cell membrane damage. Moreover, FMRPs contributed to cause stress to C. perfringens as its elongation shows. Thus, the FMRPs can be used in various food to control the pathogen. In addition, it can be expected that consumption of FMRPs may control intestinal C. perfringens as health-promoting supplements.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







