





INTRODUCTION
The crucial functions of selenium (Se) in poultry have been extensively documented. It is involved in immune responses, antioxidant defenses and other important physiological roles [1]. There are numerous reports that organic Se has greater bioavailable than inorganic Se in broilers, laying hens and quails, using Se concentrations in tissue and egg as an indicator [2,3]. Additionally, Sevcikova et al [4] found increasing Se concentrations in breast muscle and feather of broilers fed Se supplemental diets, compared to those of broilers fed the control diet. Couloigner et al [5] reported concurringly that breast muscle and feather Se concentrations were highly correlated in 21-d-old broilers. Thus, measurement of feather Se concentration could be used as a non-invasive procedure for investigation of the efficacy of Se sources for Se concentration in breast muscle. However, this suggestion should be scientifically confirmed.
Few trials have reported the responses of poultry fed supplemental organic Se from plants. Selenium from Se-enriched garlic (Allium sativum L.) and Se-enriched Chinese cabbage (Brassica pekinensis L.) was observed to improve Se status of broilers [6]. The effectiveness of Se from selenium enriched yeast (SeY) and Se-enriched mung bean sprout (Vignaradiata) was comparable in terms of egg Se concentration [7]. Dietary supplementation of Se-enriched Japanese radish sprouts (Raphanussativas) at 10 ╬╝g/kg increased serum Se level but resulted in variable Se concentrations in yolk and meat of laying hens [8]. On the other hand, Jiakui and Xiaolong [9] found the similar metabolic pathway of Se from Se-enriched malt (Hordeumvulgare L.) and sodium selenite (SS). Chantiratikul et al [10] demonstrated no different tissue Se concentrations of growing quails fed Se from SS and Se-enriched kale sprout (SeKS) cultivated in sand and watered by solution containing 60 mg Se from sodium selenate. Although, some variable outcomes had been found, those results indicated that Se from Se-enriched plants could possibly be used as an alternative Se source for poultry. Recently, hydroponically produced SeKS has been developed using SS as a Se source. Species of Se in SeKS were mostly present as organic forms [11]. The scientific reports found that the efficacy of Se from SeKS and SeY was comparable, but greater than that of Se from SS in laying quails [12], broilers [13], and laying hens [14]. Furthermore, Se from SeKS has been confirmed to be less toxic in laying hens [15] and in rats [16]. Nevertheless, there have not been any comparative studies on the effects of organic Se from SeKS and selenomethionine (SeMet) in growing quails. Therefore, the present trial aimed to compare the efficacy of dietary Se from SeKS, SeMet, and SS on productivity, carcass characteristics, meat quality, and tissue Se concentrations and to determine the relationship between Se concentrations in breast muscle and feather of growing quails.
MATERIALS AND METHODS
Animal care
The experimental procedures were approved by the Institutional Animal Care and Use Committee, Mahasarakham University (Approval No. IACUC-MSU-021/2019).
The preparation of selenium-enriched kale sprouts
After being submerged in tap water for 15 h, the kale seeds (Brassica oleracea var. alboglabra L.) were planted into sponges and placed in plastic pots. The pots were completely covered for 3 d. Afterwards, the germinated seeds were illuminated with the light from a fluorescent lamp (36 W) and watered daily. The germinated seeds were then cultivated in HoaglandŌĆÖs solution containing 30 mg Se from SS/L under the hydroponic system for 15 d. Finally, SeKS were gathered, washed, dried, ground, and stored [11].
Dietary treatments
Three hundred and twenty, 1-d-old unsexed Japanese quails (Coturnixcoturnix japonica) were obtained from a commercial farm. The quails were fed the control diet for a week. On d 7, quails were randomly divided by their body weight (26.21 ┬▒0.55 g) into four treatments. Each treatment consisted of four replicates and each replicate contained twenty quails in a Completely Randomized Design. The dietary treatments were T1: control diet; T2, T3, and T4: control diets plus 0.2 mg Se/kg from SS, SeMet (Excential Selenium4000, Orffa Additives BV, Werkendam, The Netherlands), and SeKS, respectively. Dietary inclusion level of Se at 0.2 mg/kg referred to EU regulations [17]. The control diet was prepared according to the nutrient requirement of growing quails [18], with no Se addition (Table 1). The diets were provided ad libitum to quails until slaughter at 42 d of age. Drinking water was freely available to quails at all time. The quails were reared in experimental cages (67├Ś70├Ś50 cm) placed under open housing system. Internal temperature was controlled at 28┬░C. The lighting was constantly maintained for 24 h in the cages.
Data and sample collections
Feed consumption was recorded weekly. Body weight was determined at the beginning and at the end of each period, wk 2ŌĆō3, and 4ŌĆō6. Average daily gain (ADG) and feed conversion ratio (FCR) were estimated.
On d 21, 28, 35, and 42, two birds per replicate were ran domly selected and euthanized for breast muscle (pectoralis major and minor) and wing feather collections. The samples of breast muscle and feather were dried, ground and stored at ŌłÆ20┬░C.
On d 42, blood samples of eight birds in each treatment were drawn by puncturing the wing vein. The samples were placed into sterile test tubes and centrifuged at 3,000├Śg for 10 min. The serum was harvested and stored at ŌłÆ20┬░C. Afterwards, the birds were stunned and slaughtered for breast muscle (pectoralis major and minor) collection. The fresh samples of breast muscle were stored at ŌłÆ20┬░C.
At the end of the experiment, 8 birds from each treatment were randomly weighed, stunned, slaughtered, and exsanguinated. Wings, breast muscle, legs, heart, liver, gizzard, and abdominal fat were separated and weighed for determinations of carcass characteristics. The pH values of breast muscle were measured at 45 min and 24 h post-mortem by a portable pH meter (Model HI99163, Hanna Instruments, Padova, Italy). Color measurement of breast meat was performed at 45 min and 24 h post-mortem by a Chroma meter (Model CR-410, Konica Minolta Sensing Americans, Inc., New York, USA). The breast muscle, heart, liver, and kidney were sampled, dried, and ground.
Analyses
The dietary treatment was determined for dry matter (DM, Method 934.01), crude protein (Method 976.05), ether extract (Method 920.39), crude fiber (Method 978.10), and ash (Method 942.05) according to AOAC [19]. Total Se concentrations in dietary treatments, dried tissues (breast muscle, heart, liver, and kidney), and feather were measured. Briefly, the weighed samples were wet digested by a mixture of 1.5 mL of 70% w/w nitric acid and deionized water in the digestion block at 100┬░C. When the solution had dried and cooled, 5 mL of HCl was added. The samples were then heated at 100┬░C for 10 min. After cooling the digest, deionized water was added into the volumetric flask to make up the reduced volume of the digest [20]. The Se concentrations in samples were analyzed with Atomic Absorption Spectrometer with a VGA-77 hydride generation unit (Agilent Technologies, Inc., Santa Clara, CA, USA).
Approximately 1 g of fresh breast muscle was homoge nized in 1 mL of 0.9% sodium chloride buffer using laboratory mortar with pestle. The supernatant was collected after centrifugation at 8,000 rpm for 10 min at 4┬░C and used for analysis. Glutathione peroxidase (GSH-Px) activity in breast muscle was measured according to the procedure of Paglia and Valentine [21].
Statistical analysis
All experimental data was analyzed by analysis of variance technique appropriate for Completely Randomized Design [22]. The model used was: Yij = ╬╝+Ti+╔øij, where: Yij = observation, ╬╝ = population mean, Ti = diet effect (I = 1 to 4), and ╔øij = residual error. A cage was used as the experimental unit for productive performance measurements. Selected individual quails were considered the experimental unit for carcass characteristics, GSH-Px activity and tissue Se concentration, respectively. The differences among means of each parameter were compared by DuncanŌĆÖs new multiple range test. Values of p<0.05 were considered significant. The relationships between Se concentrations in breast muscle and feather of 21 and 42-d-old quails were determined by linear regression analysis.
RESULTS
The concentrations of Se in the control diet, control diets plus 0.2 mg Se from SS, SeMet, and SeKS were 0.25, 0.46, 0.43, and 0.42 mg/kg DM, respectively (Table 2).
Effect of selenium sources on productivity and carcass characteristics of quails
The Se supplementations both in the form of inorganic (SS) and organic Se (SeMet and SeKS) did not change feed intake, ADG, FCR, carcass characteristics, and meat color of quails. However, the pH of meat at 24 h post-mortem increased (p<0.05) with Se supplementation, regardless of Se sources (Tables 3 to 5).
Effect of selenium sources on glutathione peroxidase activity and Se concentrations in tissue of quails
Dietary Se supplementation did not influence GSH-Px activity in breast muscle of quails. Breast muscle, feather, and heart tissue Se concentrations of quails fed Se from SeMet were greater (p<0.05) than those of quails fed Se from SeKS and SS. Liver and kidney tissues Se concentrations of quails fed Se from SeMet and SeKS were similar (p>0.05), but greater (p<0.05) than those of quails fed the control diet and Se from SS (Table 6).
Quails that received Se from SeMet had greater (p <0.05) breast muscle and feather Se concentrations (Tables 7 to 8) at 21, 28, 35, and 42 d of age, compared to those of quails fed Se from SeKS and SS. Selenium from SeKS increased greater (p<0.05) Se concentrations in breast muscle than Se from SS (Table 7). However, Se concentrations in feathers of quails fed Se from SeKS and SS were not different (p>0.05) as presented in Table 8.
Relationship of selenium concentrations in breast muscle and feather of quails
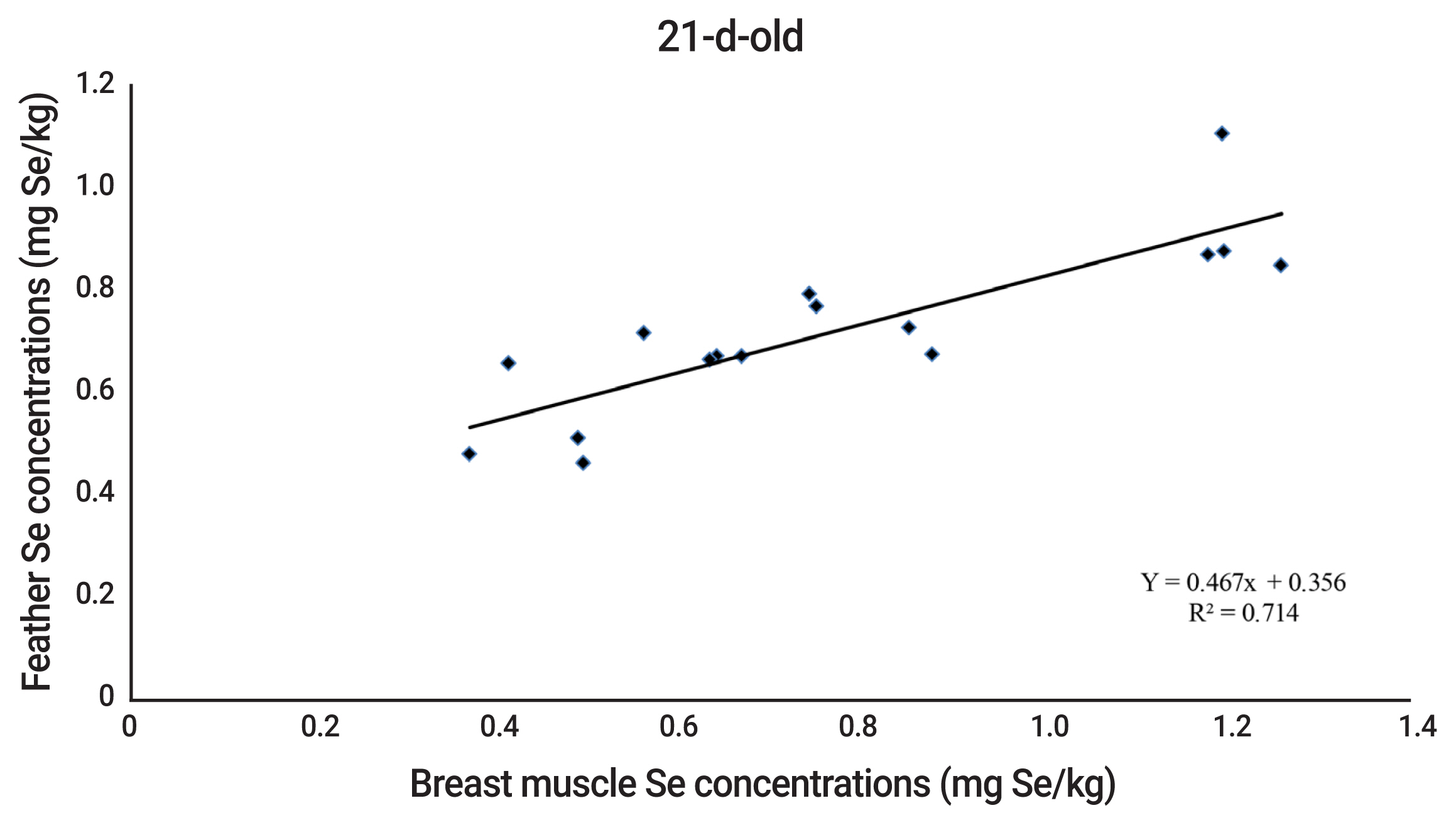
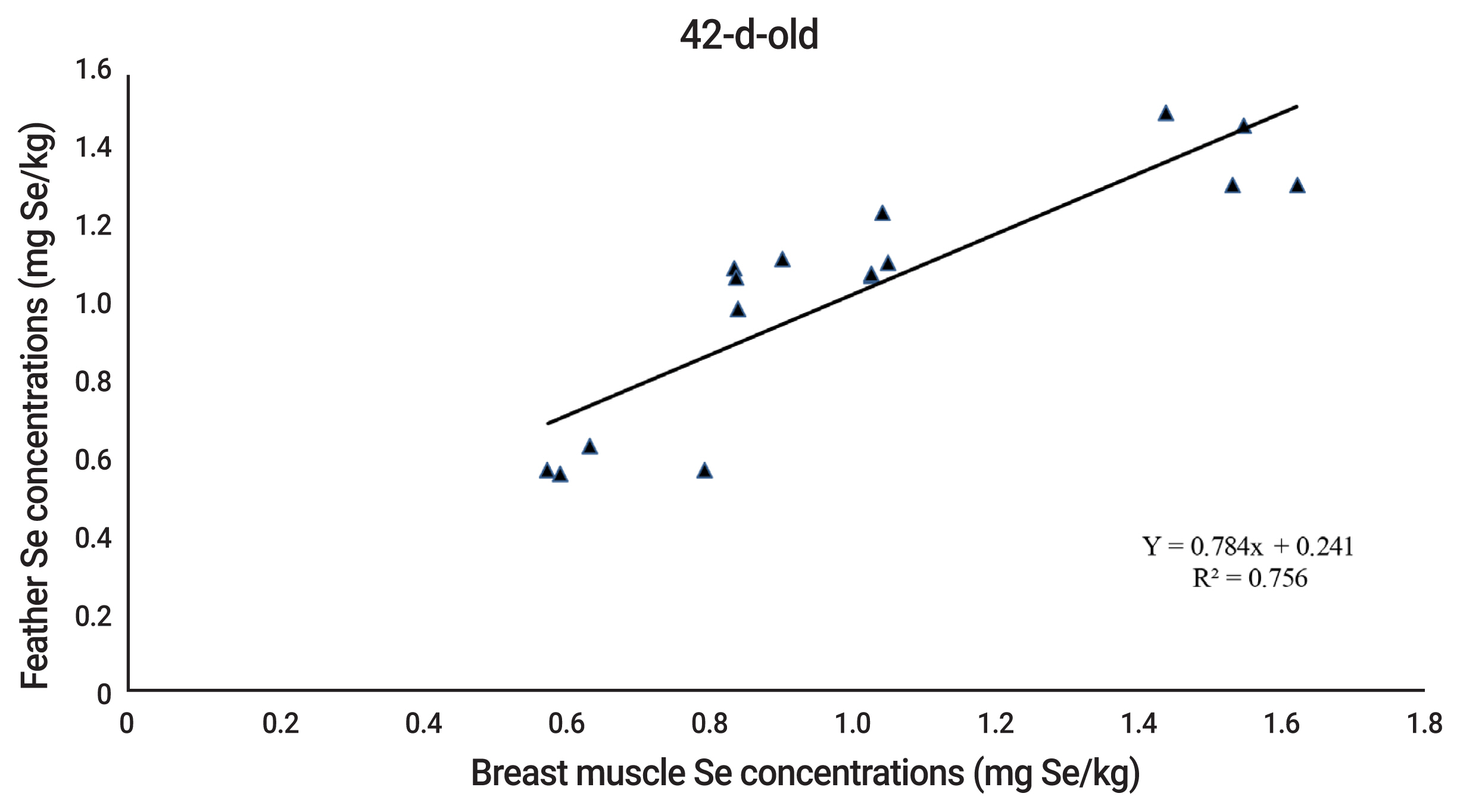
The equations of relationship between Se concentrations in breast muscle and feather of quails were Y = 0.467x+0.356 and Y = 0.784x+0.241 (where Y is breast muscle Se concentration and x is feather Se concentration) for 21 and 42 d of age, respectively. The Se concentrations in breast muscle and feather were highly correlated with R2 = 0.714, and 0.756 for 21 and 42 d of age, respectively (Figures 1 to 2).
DISCUSSION
The results demonstrated that Se supplementation in both inorganic and organic forms at 0.2 mg/kg did not change performance and carcass characteristics of growing quails. Chantiratikul et al [10] similarly found no impact of dietary Se supplementation form SeKS (0.2 to 1.0 mg/kg) on feed intake, performance, and carcass characteristics of quails. Trials in broilers also reported that Se sources from SS, SeY, SeKS, 2-hydroxy-4-methylselnobutanoic acid, and SeMet at 0.1 to 2.0 mg/kg diet did not alter feed intake and growth performance parameters [2,5], and carcass traits [13]. The Se concentration in the control diet was 0.25 mg/kg (Table 2), which met the Se requirement for quail (0.15 mg Se/kg) recommended by NRC [18]. Generally, performance of poultry reared in normal condition was not changed by supplementation of Se [6]. However, dietary Se supplementation improved productivity in heat-stressed poultry [23].
The results of this trial showed that meat color and meat pH at 45 min post-mortem were not altered by Se supplementation. However, pH of meat at 24 h post-mortem increased with Se supplementation (Table 5). Presently, there is no available published report on the effect of Se supplementation on meat quality in quail. Zhang et al [24] found that Se supplementation (0.3 mg/kg) from SS and SeMet did not improve meat color of 56-d-old offspring of broiler breeders. However, Wang et al [25] reported that broilers fed Se form SS and SeMet (0.15 mg/kg) resulted in increasing meat color value of breast muscle at 8 and 16 h post-mortem. Zhan et al [26] reported similarly that adding SS or SeMet in finishing pig diets caused an increasing trend of the pH value of loin muscle at45 min post-mortem in Se-treated groups. These results are similar to ours presented here. Generally, lower meat pH reduces the muscle protein ability to bind to water, causing shrinkage of myofibrils [26]. Therefore, the present results of the effect of Se supplementation on pH of meat at 24 h post-mortem reflected the observation that Se supplementation improved meat quality in quails. Nevertheless, additional research should be conducted for clarification of the effects of dietary Se addition on meat quality. Moreover, the above inconsistent findings might be due to different levels of Se supplementation, Se sources, Se species and animals used in the experiment.
GSH-Px is a selenoprotein enzyme, containing Se. It is an important antioxidant enzyme in animals. Our results found an insignificant effect of both inorganic (SS) and organic Se (SeMet and SeKS) supplementation on GSH-Px activity in breast muscle of quails (Table 6). Wang et al [25] have found similarly that GSH-Px activity in breast muscle of broilers was not affected by inorganic or organic Se supplementation. Those results could be explained by metabolic pathway of dietary Se. The metabolic route of Se is illustrated that both inorganic and organic Se can be effectively converted to hydrogen selenide (H2Se), which is used for selenoproteins synthesis such as GSH-Px [27].
The findings indicated that organic Se from SeMet and SeKS effectively increased Se concentrations in tissues of quails, compared to inorganic Se from SS (Table 6). Numerous reports have also demonstrated that organic Se has a better accumulative efficacy in tissues of poultry than inorganic Se [5,13,24]. The Se concentrations in breast muscle, heart tissue and feather of quails fed SeMet were significantly greater than those of quails fed SeKS. These results probably reflected a better efficiency of Se from SeMet compared to Se from SeKS. Additionally, breast muscle and feather Se concentrations (Tables 7 to 8) of quails at 21, 28, 35, and 42 d of age clearly confirmed the greater efficiency of Se from SeMet than those of SeKS. SeMet has been previously reported regarding its greater efficacy on Se depositions in tissues of broilers and in egg of laying hens than those of SeY [1]. The current trial evaluated the efficiency of Se from SeMet and SeKS using Se concentrations in tissues of growing quails as an index. The bioavailability of Se in animals mainly depends on SeMet content. SeMet can be built into body protein instead of methionine and allows Se accumulation in the tissues, resulting in greater Se concentration in muscle and other tissues [28]. The SeMet used in this trial contains a constant level of L-SeMet [28]. However, SeKS consists of SeMet and methyselenocysteine at 41% and 35%, respectively [11]. The greater SeMet content in SeMet than that in SeKS resulted in greater Se concentrations in tissues and feather of quails. The obtained results confirmed that Se from SeMet was more efficiently deposited in tissues of quails compared to Se from SeKS. The Se from SS had lower efficacy in tissue accumulation than Se from SeMet and SeKS. Additionally, kidney Se concentrations of quails fed both organic and inorganic Se seemed to be greater than other tissues Se concentrations (Table 6). Those results agree with the previous research in laying quails [12]. Ingested Se is generally used for selenoproteins synthesis after conversion to H2Se. However, unmetabolized Se will be mainly excreted via urine [27], resulting in greater Se accumulation in kidney than other tissues.
Naturally, pre-juvenile molting begins when the quails are 3-d-old. Juvenile body plumage is complete in about 30 days [29]. During this period, the second generation of feathers will be developed to replace the down feathers. Thus, Se content in the second generation of feathers depends on Se source and level in the diet. There is little information concerning relationship between Se in muscle or other tissues and feather of poultry. Quails fed drinking water with added SS or SeMet had the highest Se concentrations in the pancreas, followed by the down feathers, liver, and kidney [30]. Sevcikova et al [4] observed that Se concentrations in breast muscle and feather of 35-d-old broilers increased accordingly with dietary Se supplementation. Similarly, other researchers reported that the correlation coefficient between muscle and feather Se concentrations of 21 d-old broilers was 0.927 [5]. The present results concurringly found that Se concentrations in breast muscle and feather of 21 and 42-d-old quails were strongly related (R2 = 0.714 and 0.756 for 21 and 42-d-old, respectively). The results, therefore, reflect that feather Se concentration, instead of muscle Se concentration, can be used as an indicator for assessment of Se efficacy of Se sources in 21 and 42-d-old quails.
In conclusion, productive performance, carcass charac teristics, meat color, and GSH-Px activity in breast muscle of quails were not affected by dietary Se supplementation. Selenium from SeMet was more effective in increasing Se concentrations in the tissues of quails than Se from SeKS and SS. Feather Se concentrations of 21 and 42-d-old quails can be used for assessment of Se bioavailability of dietary Se sources.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







