





INTRODUCTION
A disintegrin and metalloproteinase with thrombospondin motifs (ADAMTSs) are secreted, extracellular enzymes with a compound domain organization [1]. In humans, nineteen ADAMTS proteins are encoded in the genome and they can be clustered into eight ŌĆścladesŌĆÖ based on their domain organization and known functions. The aggrecanase and proteoglycanase clades (ADAMTS1, 4, 5, 8, 9, 15, and 20) can cleave hyaluronan-binding chondroitin sulfate proteoglycans, a type of extracellular proteins, including aggrecan, versican, brevican, and neurocan [2]. Among them, ADAMTS8, also known as metalloprotease and thrombospondin domains 2 (METH-2), is a member of the ADAMTS family and was originally identified as an antiangiogenic factors that is down-regulated in brain cancer [3,4]. ADAMTS8 down-regulation has been also reported in other cancers, including brain, breast, lung, pancreatic, and thyroid cancer [4ŌĆō7]. The ADAMTS8 gene shows a high frequency of promoter methylation in brain, lung, and thyroid cancer, suggesting that the epigenetic silencing of ADAMTS8 may be involved in tumorigenesis [4,6,8]. Moreover, in cancer cells regulated by ADAMTS8, certain mutations can drive abnormal signaling during viral infection, indicating a correlation between cancers and viruses [9,10].
Pathogen-associated molecular patterns (PAMPs), such as polyinosinic-polycytidylic acid (poly[I:C]) and lipopolysaccharide, have been used to study toll-like receptor (TLR)-mediated cellular responses. Poly(I:C) is a synthetic analogue of double-stranded RNA (dsRNA), a PAMP generated during the replication of RNA and DNA viruses [11], and is recognized by distinct receptors depending on their localization. When added to the culture medium, poly(I:C) is mainly sensed by endosome-localized TLR3 [12,13]. TLR3 signaling can also occur in non-immune cells, contributing to an antitumor response. TLR3 is activated by extracellular dsRNA, which is recognized by the receptor in a sequence-independent manner. TLR3 initiates a protective response against dsRNA viruses including polio virus, coxsackievirus group B and serotype 3, and encephalomyocarditis virus, as well as DNA viruses, such as herpes simplex virus 1 and murine cytomegalovirus [14ŌĆō16]. Besides the mammals, TLR3 recognizes dsRNA virus infection and is involved in the resistance or susceptibility to viral infection in fowls. It was reported that the mRNA-expression levels of duck TLR3 and other cytokines (including interferon-╬▒ [IFN-╬▒]) were highly up-regulated during infection by duck reovirus, a dsRNA virus [17]. In 2014, Cheng et al [18] reported that infection with the chicken Newcastle disease virus increased the mRNA expression of chicken TLR3. ChTLR3 actively participates in the recognition of pro-inflammatory responses during viral infection, and leads to consequent antiviral cytokine secretion in chickens. In addition, TLR3 recognizes dsRNA that has been transcribed in vitro and its synthetic analogues, such as poly(I:C) and polyadenylic:polyuridylic acid (poly [A:U]). These analogues have been used to mimic responses to RNA virus infection and are commonly administered in in vitro and in vivo studies of TLR3-mediated cellular responses [19].
Although the expression of chicken ADAMTS8 (chADAMTS8) appears to be related to virus infection, the structure and expression pattern of this gene have not been studied. Here, we analyzed the amino acid sequence encoded by the chADAMTS8 gene by comparing it with the ADAMTS8 amino acid sequences from other species. Gene expression was investigated in various chicken tissues. In addition, the expression pattern of the chADAMTS8 gene was evaluated in the chicken DF-1 fibroblast cell line after stimulation with poly(I:C).
MATERIALS AND METHODS
Cell culture and stimulation
The chicken DF-1 cell line was purchased from the American Tissue Culture Collection (CRL-12203, Manassas, VA, USA). DF-1 cells were cultured in DulbeccoŌĆÖs modified EagleŌĆÖs medium supplemented with 10% fetal bovine serum, 2 mM L-glutamine, and 100 U/mL each of penicillin and streptomycin (Thermo Scientific, Logan, UT, USA) at 37┬░C in a humidified atmosphere of 5% CO2 and 95% air. Poly(I:C) was purchased from Invivogen (San Diego, CA, USA) and used for stimulation at a concentration of 10 ╬╝g/mL for 24 h.
Bioinformatic analysis
The ADAMTS8 mRNA and amino acid sequences of various species (chicken, duck, human, chimpanzee, mouse, rat, cow, pig, and horse) were retrieved from the Ensembl database (http://www.ensembl.org/) (Table 1) and aligned with the BioEdit software, using the ClustalW method. The protein domains were predicted by using the SMART domain search program (http://smart.embl-heidelberg.de/). Phylogenetic analyses were performed with the MEGA7 software [20].
Reverse transcription-polymerase chain reaction
Trizol (Invitrogen, Carlsbad, CA, USA) was used to extract total RNA from tissue samples (liver, lung, kidney, spleen, and heart) and DF-1 cells. Total RNA was quantified using a NanoDrop spectrophotometer (Thermo Fisher Scientific Inc., Waltham, MA, USA). The chADAMTS8, chTLR3, and chTLR4 transcripts were analyzed by reverse transcription-polymerase chain reaction (RT-PCR) amplification. The RT-PCR conditions were as follows: an initial step of 94┬░C for 10 min; 35 cycles of 94┬░C for 30 s, 60┬░C for 30 s, and 72┬░C for 30 s; and a final step of 72┬░C for 10 min. The RT-PCR products were analyzed by electrophoresis on a 2.0% SeaKem LE agarose gel (Lonza, Basel, Switzerland). Target gene expression was normalized against that of the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene. The sequences of the GAPDH primers were 5ŌĆ▓-TGC TGC CCA GAA CAT CAT CC-3ŌĆ▓ for the forward primer and 5ŌĆ▓-ACG GCA GGT CAG GTC AAC AA-3ŌĆ▓ for the reverse primer.
Quantitative real-time polymerase chain reaction analysis
Quantitative RT-PCR (qRT-PCR) was conducted with a CFX-96 RT-PCR detection system (BioRad, Hercules, CA, USA) to analyze chADAMTS8, chTLR3, chTLR4, and chIL1B expression. The sequences of the chADAMTS8 primers were 5ŌĆ▓-GCA CTA TGA CAC TGC CAT CCT-3ŌĆ▓ for the forward primer and 5ŌĆ▓-CGT GTC GCA GCC TTG ATG-3ŌĆ▓ for the reverse primer. ChTLR3 primers were: 5ŌĆ▓-CCA TTT TGA AGG GTG GAG AA-3ŌĆ▓ (forward) and 5ŌĆ▓-CCT GCT TCG AAG TCT CGT TC-3ŌĆ▓ (reverse). chTLR4 primers were: 5ŌĆ▓-TTC CAA GCA CCA GAT AGC AAC ATC-3ŌĆ▓ (forward) and 5ŌĆ▓-ACG GGT CAC AGA AGA ACT TAG GG-3ŌĆ▓ (reverse). chIL1B primers were: 5ŌĆ▓-GGA TTC TGA GCA CAC CAC AGT-3ŌĆ▓ (forward) and 5ŌĆ▓-TCT GGT TGA TGT CGA AGA TGT C-3ŌĆ▓ (reverse). The PCR conditions were as follows: an initial step of 94┬░C for 3 min; 39 cycles of 94┬░C for 10 s, 60┬░C for 30 s, and 72┬░C for 30 s; and a final step of 72┬░C for 10 min. Dissociation was performed at 0.5┬░C increments from 55┬░C to 95┬░C over 25 min. All samples were measured in triplicate to ensure reproducibility, and Ct values were calculated by the 2ŌłÆ╬ö╬öCt method [21]. Expression of the GAPDH gene was detected as the reference.
Statistical analysis
Results are presented as the means┬▒standard deviation of triplicate independent experiments. Statistical significance was assessed using a StudentŌĆÖs t-test. A p value of <0.05, compared with the non-treated control, was considered to reflect a statistically significant difference.
RESULTS AND DISCUSSION
The amino acid sequence encoded by chADAMTS8 was analyzed and compared with that of the other species. ChADAMTS8 was identified as a differentially expressed gene (DEG) from chicken kidney by RNA-Seq analysis, after the chickens were fed a diet containing different amounts of calcium [22]. The chADAMTS8 amino acid sequence was compared with the duck, horse, pig, cow, mouse, rat, chimpanzee, and human sequences, which were retrieved from the Ensembl database. ChADAMTS8 was predicted to contain three thrombospondin type 1 (TSP1) domains, whose amino acid sequences were conserved among the species examined, whereas sequences outside of these domains (especially in the amino-terminal region) were diverse (Figure 1A). Specifically, all three TSP1 domains of duck ADAMTS8 shared homology with chADAMTS8, whereas only two TSP1 domains were homologous with those of the other mammal species. Nonetheless, this result suggests that the chADAMTS8 may serve similar biological functions as ADAMTS8 in other species, including antiangiogenic activity and tumorigenesis. The phylogenetic tree of ADAMTS8 revealed that chADAMTS8 is evolutionarily clustered in the same clade with that of the duck (Figure 1B).
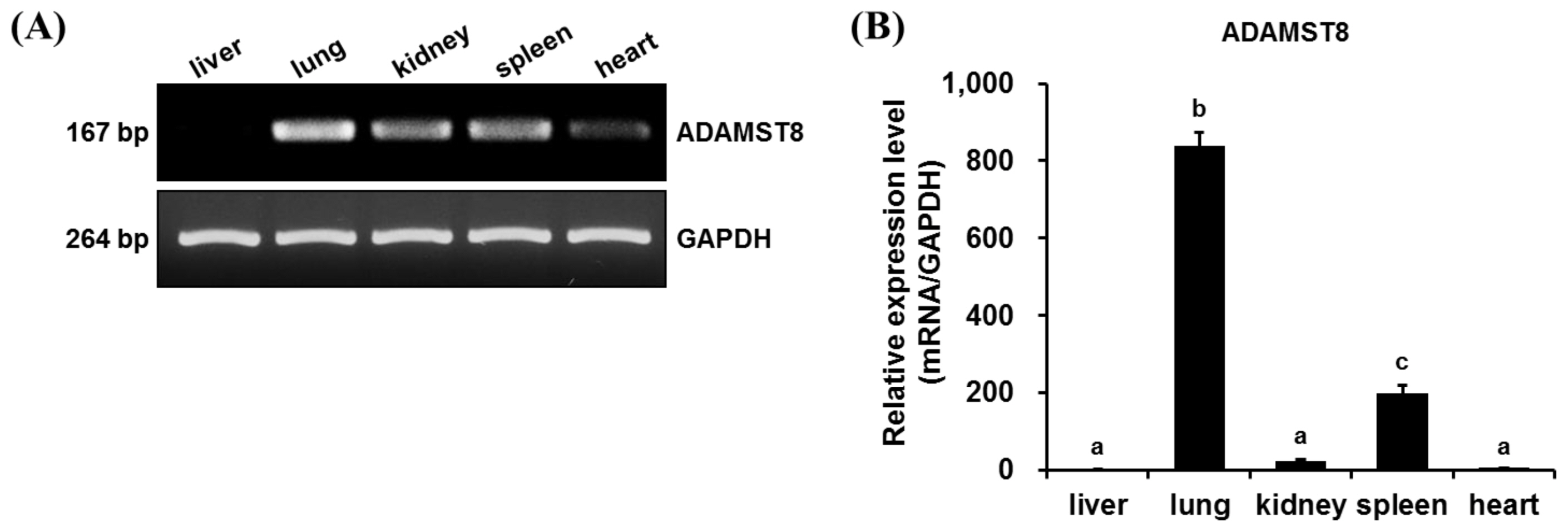
ADAMTS8 is known to be related to the development of lung cancer and recently, Zhou et al [23] reported that ADAMTS8 can be used as a biomarker for human lung cancer [6]. The gene-expression pattern of chADAMTS8 was investigated in various chicken tissues. RT-PCR and qRT-PCR analysis revealed that chADAMTS8 gene was highly expressed in the chicken lung tissue under normal conditions (Figure 2A, 2B). In addition to lung tissue, chADAMTS8 was also expressed in other tissues including spleen, kidney, heart, and liver. Like human ADAMTS8 gene expression in normal adult and fetal lung tissues, chADAMTS8 showed the highest expression in lung tissue and the second-highest expression in the spleen tissue. So far, it is not clear what the physiological role of chADAMTS8 is even though its expression is the highest in lung, and further study warrants revealing its role in normal condition as well as inflammatory conditions, caused by viral pathogens.
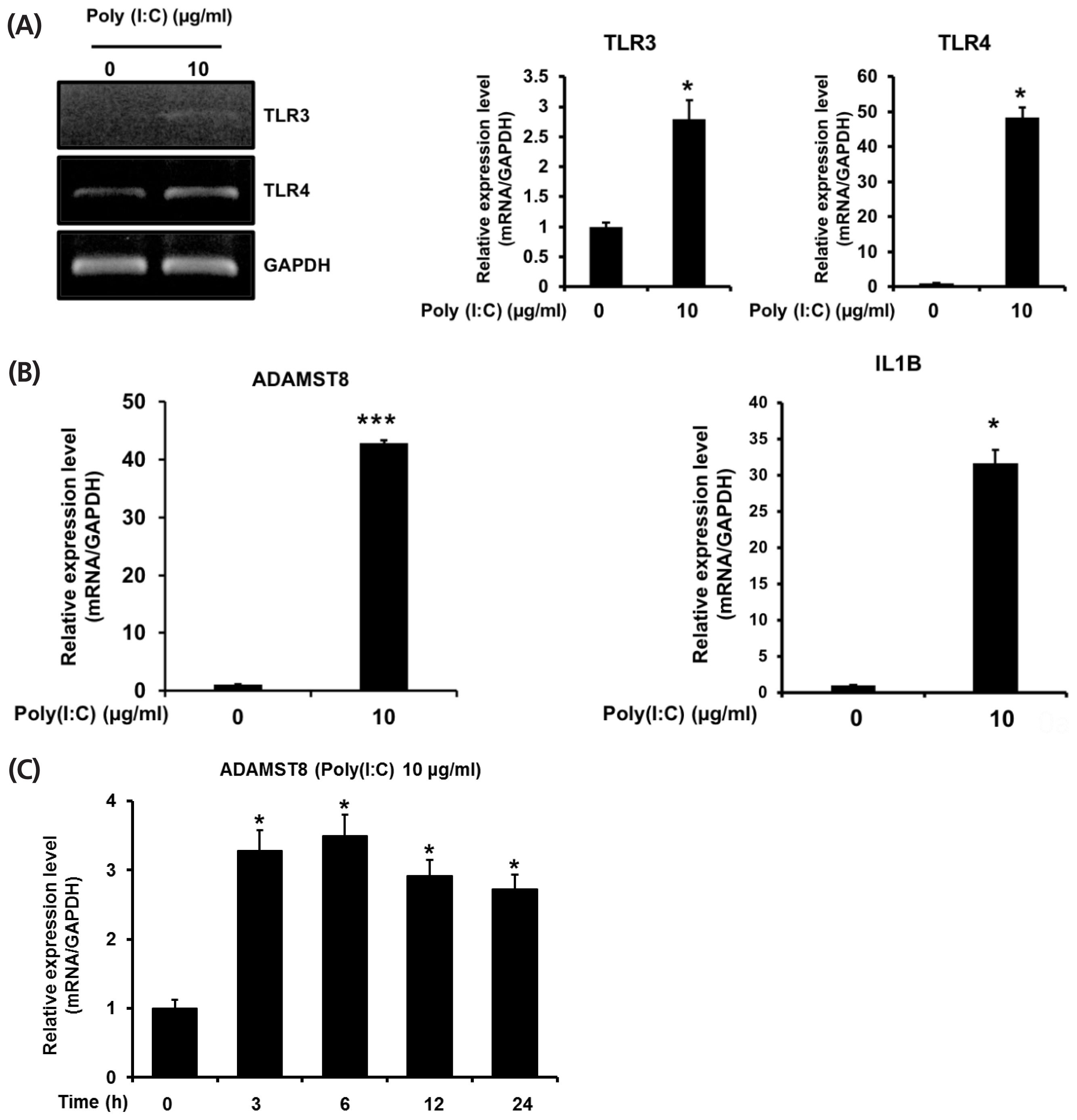
In addition, to gain insight into the regulation of chADAMTS8 gene expression, we examined its expression in chicken DF1 cells that were stimulated with poly(I:C). To confirm the innate immune responses of chicken DF-1 cell is stimulated by poly(I:C), the activation of TLR3 and TLR4 was examined with the agonist treatment. As shown in Figure 3A, the expressional levels of TLR3 and TLR4 in chicken DF-1 cell were increased under stimulation of poly(I:C). Additionally, the downstream activation of TLR3 pathway in response to the poly(I:C) stimulation was observed with interleukin 1 beta expression which is a cytokine induced following TLR3 activation (Figure 3B). A recent study reported that DF-1 cells express higher level of suppressor of cytokine signaling 1 (chSOCS1), a negative regulator of cytokine signaling in mammals, than chicken embryonic fibroblast cells, suggesting the dampened signaling activity of DF-1 cells in IFN-alpha (IFN-╬▒) signaling pathways through SOCS1 by inhibiting Janus kinase (JAK)-signal transducers and activator of transcription (STAT) signaling axes [24]. Nonetheless, our results show that chTLR3, chTLR4, and chIL1b expressions, as TLR3 signal target genes, are induced in poly(I:C) stimulation, suggesting that TLR3 signaling pathway of DF-1 cells may be separate from JAK-STAT signaling pathway activated by IFN-╬▒,╬▓, although further study is required to address it. Figure 3C shows qRT-PCR results of ADAMTS8 expression in chicken DF-1 cells with the treatment of poly(I:C). After TLR3/Mda5 agonist poly(I:C) stimulation, ADAMTS8 expression was measured time-dependently at 3, 6, 12, and 24 h. The expressional level of chADAMTS8 was increased as early as 3 h, and after that, the level was not elevated further until 24 h. Overall, these results suggest that chADAMTS8 expression may respond specifically to viral infection and thus, be correlated with TLR3-mediated cellular responses.
The complete molecular characterization of ADAMTS8 will be challenging; however, the genetic information and expression patterns of chADAMTS8 presented here can provide the foundation for further study including the transcriptional inhibition effects on the chADAMTS8 expression under the agonists poly(I:C). In addition, the up-regulation of chADAMTS8 after poly(I:C) treatment suggests that this gene might be related to viral infection in chickens and that chADAMTS8 gene is a promising candidate biomarker for infectious diseases that are rampant in chickens.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







