




INTRODUCTION
Feedlot cattle industry in the United States uses ionophores as feed additive which act in one or more ways in the animal like improve weight gain, feed efficiency, reduce morbidity, mortality and may act favorably on environment. The action mode of ionophores is well established, they act against gram positive bacteria, improve propionate, reduce acetate, also reduce deamination, methane, risk of bloat and lactic acidosis. As an alternative to ionophores, virginiamycin (VM) is an antimicrobial also effective against growth of gram-positive bacteria, including ruminal lactic acid-producing bacteria, reducing the possibility of lactic acidosis associated digestive dysfunctions. In a summary of 7 dose-response studies with feedlot cattle [1], dietary supplementation with 19 to 27 mg VM/kg did not affect dry matter intake (DMI), but enhanced average daily gain (ADG, 4.6%) and gain efficiency (ADG/DMI, 3.6%), and reduced (38%) the incidence of liver abscess. In more recent studies involving cross-bred [2] and calf-fed Holstein steers [3,4] VM supple-mentation likewise enhanced ADG (6.1%) and gain efficiency (7.2%). In these latter studies, it was shown that the improvement in gain efficiency with VM supplementation was largely due to enhanced efficiency of energy utilization, increasing the net energy value of the diet for maintenance (NEm, 6.6%) and gain (NEg, 7.9%). The VM supplemen-tation appears to have very little impact on the extent of organic matter (OM) digestion, and the digestible energy value of the diet, per se [3]. The VM decreases ruminal deaminase activity and ruminal degradation of feed protein. Montano et al [2] observed that VM supplementation increased flow of feed N to the small intestine. No research has been reported thus far that evaluates the potential for a differential response to VM supplementation depending on the energy density of the finishing diet. To the extent that the growth enhancement response to VM supplementation is dependent on an amelioration of episodic digestive dysfunctions associated with prolonged feeding of high-grain finishing diets, it follows that the response to VM supplementation might diminish as the grain content of the diet is reduced. The objective of the present study was to examine the differential response to VM supplementation on growth-performance and characteristics of digestion when the grain content of the diet is reduced by 25%, decreasing dietary NEm from 2.22 to 2.10 Mcal/kg.
MATERIALS AND METHODS
All procedures involving animal care and management were in accordance with and approved by the University of California, Davis, Animal Use and Care Committee.
Trial 1. Growth performance and carcass characteristics of feedlot steers
The magnitude of ionophore antibiotic response can be greatly influenced by energy density of the diet. Eighty crossbred beef steers (298.2Âą6.3 kg) were used in a 152-d experiment to compare the effects of energy density and VM supplementation on growth performance, dietary net energy (NE), and carcass characteristics. Cattle originated from southeast Texas and were received at the University of California Desert Research Center, El Centro, on April 3, 2013. Upon arrival, steers were vaccinated for bovine rhinotracheitis-parainfluenza (Bovi-Shield Gold, Zoetis, New York, NY, USA), clostridials (Ultrachoice-7, Zoetis, New York, NY, USA), treated for parasites (Dectomax Injectable, Zoetis, USA), injected subcutaneously with 500,000 IU vitamin A (Vital E-A+D3, Stuart Products, Bedford, TX, USA), and 800 mg tulathriomycin (Draxxin, Zoetis, USA), branded, ear-tagged, and implanted with Revalor-S (Intervet, Millsboro, DE, USA). Horns, if present, were tipped. Steers were then blocked by weight and randomly assigned within weight groupings to 16 pens, 5 steers/pen. Pens were 78 m2 with 33 m2 of overhead shade, automatic waterers, and fence-line feed bunks. Composition of experimental diets is shown in Table 1. The 152-d trial consisted of a 28-d receiving period followed by a 124-d growing finishing period. During the receiving period steers were fed a receiving diet for 14 d, followed by 7 d each on two successive transition diets. Thereafter, steers were fed either a lesser energy dense (LED, 2.10 Mcal/kg NEm) or higher energy dense (HED, 2.22 Mcal/kg NEm) growing finishing diet. All diets were fed with or without 28 mg/kg (dry matter [DM] basis) VM (V-max 50, Phibro Animal Health, Ridgefield Park, NJ, USA) in a 2Ă2 factorial arrangement. Diets were prepared at weekly intervals and stored in plywood boxes located in front of each pen. Weekly samples of each diet were composited at 28-d intervals for dry matter determination. Steers were allowed ad libitum access to their experimental diets. Fresh feed was provided twice daily. On d 84 of the trial, all steers were injected subcutaneously with 500,000 IU vitamin A (Vital E-A+D3, Stuart Products, Bedford, TX, USA) and re-implanted with Revalor-S (Intervet, USA). Two steers, randomly selected from each pen, were orally bolused with a ruminal data logger (IDL-750, Telonics, Mesa, AZ, USA) for continuous recording of ruminal temperature at 15 min intervals.
Individual live weight (LW) and feed intake were determined at 28-d intervals. Feed intake was determined as the amount of feed offered minus the residual feed remaining in the feed bunk at the time cattle were weighed (a few h prior to the morning feeding). For calculating steer performance, initial LW is the off-truck arrival weight. Interim and final LW were reduced 4% to account for digestive tract fill. Final shrunk LW was adjusted for hot carcass weights (HCW) by dividing HCW by the decimal fraction of the average dressing percentage (0.632). Gain efficiency is ADG/DMI. Energy gain (EG) was calculated by the equation: EG = ADG1.097(0.0557[WĂ478/527]0.75), where EG is the energy deposited (Mcal/d), W is the average shrunk body weight in kg, and 527 is the average harvest weight [5]. Maintenance energy (EM) was calculated by the equation: EM = 0.077W0.75 [6]. Dietary NEg was derived from NEm by the equation: NEg = 0.877NEmâ0.41. The DMI is related to energy requirements and dietary NEm according to the equation: DMI = EG/(0.877NEm â0.41)+EM/NEm, and can be resolved for estimation of dietary NE by means of the quadratic formula:
x = ( - b - b 2 - 4 a c ) / 2 c
The HCW were obtained at time of slaughter. After carcasses chilled for 48 h, the following measurements were obtained: longissimus muscle (LM) area (cm2) by direct grid reading of the LM at the 12th rib; subcutaneous fat (cm) over the LM at the 12th rib taken at a location 3/4 the lateral length from the chine bone end (adjusted by eye for unusual fat distribution); kidney, pelvic, and heart fat (KPH) as a percentage of HCW; marbling score [8]; using 3.0 as minimum slight, 4.0 as minimum small, 5.0 as minimum modest, 6.0 as minimum moderate, etc.), and estimated retail yield of boneless, closely trimmed retail cuts from the round, loin, rib, and chuck (% of HCW; [9]) = 52.56â1.95Ăsubcutaneous fatâ1.06ĂKPH+0.106ĂLM areaâ0.018ĂHCW.
Pens were used as experimental units. In addition to overall 152-d treatment effects on cattle growth performance, treatment effects were further separated into initial 84-d growing phase (representing the period of first growth implant), and 68-d finishing phase (representing the period of high ambient temperature following application of the second growth implant and insertion of ruminal thermistors). The experimental data were analyzed as a randomized complete block design in a 2Ă2 factorial arrangement (Statistix 10, Analytical Software, Tallahassee, FL, USA). Treatment effects were considered significant for pâ¤0.05 and as trends for p>0.05 and â¤0.10. When significant interactions were detected, mean separation (least significant difference, LSD) was performed.
Trial 2. Characteristics of digestion and ruminal fermentation
Four Holstein steers (170.4Âą5.6 kg) with cannulas in the rumen (3.8 cm internal diameter) and proximal duodenum [10] were used in 4Ă4 Latin square experiment to study treatment effects on characteristics of digestion. Treatments were the same as those used in Trial 1 (Table 1) with the inclusion of 0.30% chromic oxide as a digesta marker. Dietary VM was top-dressed on feed allotment at time of feeding. Steers were maintained in individual pens (4 m2) with automatic waterers. Diets were fed at 0800 and 2000 h daily. In order to avoid the complications of feed refusals, DMI was restricted to 3.72 kg/d (equivalent to 2.2% of LW). Experimental periods were 3 weeks, with 10 d for dietary treatment adjustment, 4 d for collection, and 7 d of drug withdrawal (steers were fed only the basal diet with no added VM for 1 week before switching to new dietary treatment assignments of the subsequent experimental period). During collection, duodenal and fecal samples were taken twice daily as follows: d 1, 0750 and 1350 h; d 2, 0900 and 1500 h; d 3, 1050 and 1650 h, and d 4, 1200 and 1800 h. Individual samples consisted of approximately 700 mL of duodenal chime and 200 g (wet basis) of fecal material. Samples from each steer within each collection period were composited for analysis. During the final day of each collection period, ruminal samples were obtained from each steer via ruminal cannula 4 h after feeding. Ruminal fluid pH was determined on fresh samples. Samples were strained through 4 layers of cheesecloth. Two milliliters of freshly prepared 25% (wt/vol) meta-phosphoric acid was added to 8 mL of strained ruminal fluid. Samples were then centrifuged (17,000Ăg for 10 min), and supernatant fluid was stored at â20°C for volatile fatty acids (VFA) analysis (gas chromatography) [11]. Upon completion of the experiment, ruminal fluid was obtained via the ruminal cannula from all steers and composited for isolation of ruminal bacteria by differential centrifugation [12].
Samples were subjected to all or part of the following analysis: dry matter (oven drying at 105°C until no further weight loss), ash, ammonia nitrogen (method 8.026; [13]), Kjeldahl nitrogen (method 7.037; [13]), neutral detergent fiber ([14]; adjusted for insoluble ash), chromic oxide [15], gross energy (adiabatic bomb calorimetry), purines [16], and starch [17]. Microbial OM and nitrogen leaving the abomasum were calculated using purines as a microbial marker [16]. The OM fermented in the rumen was considered equal to OM intake minus the difference between the amount of total OM reaching the duodenum and microbial OM reaching the duodenum. Feed N escape to the small intestine was considered equal to total N leaving the abomasum minus ammonia-N, microbial N and endogenous N, assuming endogenous N is equivalent to 0.195W0.75 [18]. Methane production (mol/mol glucose equivalent fermented) was estimated based on the theoretical fermentation balance for observed molar proportion of acetate, propionate, and butyrate (Ma, Mp, and Mb, respectively), where mol methane, Mm = Ma+2Mbâ(Ma/2+ Mp/4+3Mb/2), and mol glucose equivalent = (2Ma+3Mp+4Mb+ (Ma/2+Mp/4+3Mb/2)+ Mm)/6 [19]. Digestible energy was calculated as gross energy intake minus gross energy excretion.
Treatment effects on characteristics of digestion were ana lyzed as a 4Ă4 Latin square design with 2Ă2 factorial arrangement of treatments (Stastix 10, Analytical Software, USA). The statistical model for the trial was as follows: Yijk = Îź+Si+Pj+Tk+Eijk, where: Yijk is the response variable, Îź is the common experimental effect, Si is the steer effect (df = 3), Pj is the period effect (df = 3), Tk is the treatment effect (df = 3) and Eijk is the residual error (df = 6). Treatment effects were considered significant for pâ¤0.05 and as trends for p>0.05 and â¤0.10. When significant interactions were detected, mean separation (LSD) was performed.
RESULTS AND DISCUSSION
Trial 1. Growth performance and carcass characteristics of feedlot steers
Treatment effects on cattle growth performance are shown in Table 2. The VM supplementation did not influence on ADG (p>0.10). Based on growth equations of Zinn et al [20]; ADG = [1.628+0.00287IWâ0.00000107 IW2â0.461 Frame]), expected ADG for medium-frame steers in this study (initial weight [IW] = 298.2 kg) is 1.47 kg, in good agreement with the observed ADG (1.50 kg). Likewise, Salinas-Chavira et al [3] did not observe an effect of VM supplementation on 340-d ADG of calf-fed Holstein steers. Nevertheless, in other studies [1,2,4], increased ADG has been a distinctive response to VM supplementation.
During the initial 84-d period, LED-fed steers tended to have greater (7.4%, p = 0.08) ADG than HED-fed steers. However, during the subsequent 68-d period and overall, the effect of diet energy density on ADG was not appreciable (p>0.10). This effect was expected, as energy density of both the LED and HED diets were sufficiently high so to not limit energy intake, per se [5,21].
Daily DMI was greater (p <0.01) for LED- than for HED-fed steers, resulting in decreased gain efficiency. These differences were anticipated. Indeed, observed DMI was in good agreement (98%) with expected based on ADG and diet energy density. However, there was a tendency (p = 0.09) for an interaction between VM supplementation and diet energy density on DMI. With the LED diet VM tended to increase DMI, whereas with the HED diet VM tended to decrease DMI. In the case of the LED diet, the tendency for increased DMI is reflected in a numerical increase in ADG. Supplemental VM did not affect (p = 0.34) the estimated NE value of the LED diet. In contrast, in the case of the HED diet, the tendency for decreased DMI with VM supplementation was due to an effectual increase of 5% and 7%, respectively (p<0.05), in estimated dietary NEm and NEg. The magnitude of the increase in dietary NEm and NEg is in good agreement with previously reported increases of 6% and 7%, respectively [2â4]. There were no treatment effects on carcass characteristics (p>0.10; Table 3).
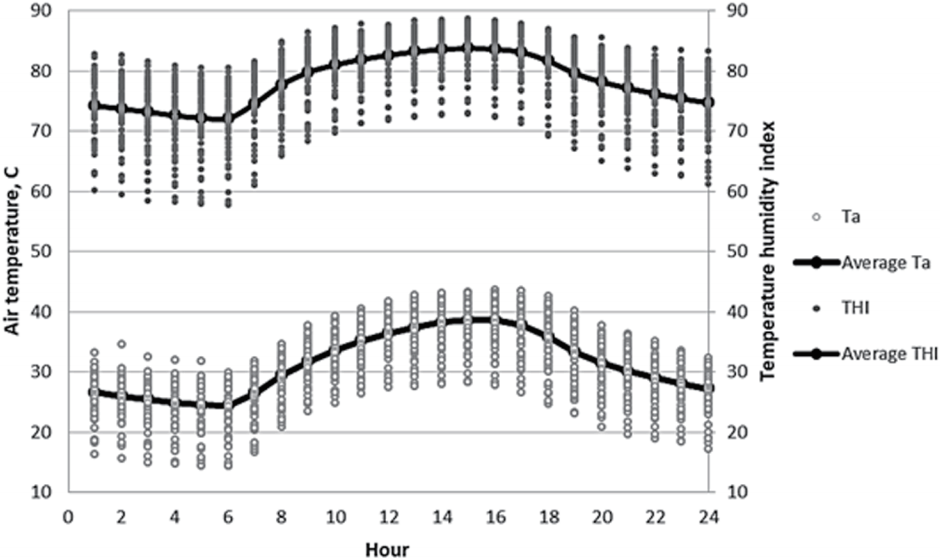
Hourly air temperature (Ta), temperature humidity index (THI [22]; NOAA [23]), and ruminal temperature during a 70-d period from July 25 through October 3, 2013 are shown in Figure 1, 2. There was a reasonably close association between both average Ta and THI and ruminal temperature, explaining 72.4% and 66.9% of the variation, respectively (p<0.01). Mader et al [24] observed that under standard conditions of solar radiation and wind, day-time respiration rate of feedlot cattle (Angus and Angus cross) was classified as âelevatedâ when THI approached 70 and âmoderateâ (including presence of small amount of drool or saliva) when THI approached 80. During the 70-d monitoring period of the present study, 24-h THI averaged 78.2Âą6.2. Hourly THI averaged 80 or greater between the period 0900 and 1900 h. During the time period between 1900 and 0700 h (evening and early morning), there were no treatment effects (p>0.10) on ruminal temperature. During the daylight hours between 0700 and 1900 h, diet energy density did not affect (p = 0.97) ruminal temperature, averaging 40.35°C and 40.36°CÂą0.15°C for the LED and HED diets, respectively. However, during that period VM supplementation decreased (p<0.05) ruminal temperature, averaging 40.50°C and 40.21°CÂą0.05°C for non-supplemented and VM supplemented diets, respectively. In broiler chickens under condition of heat stress, enhanced growth performance with VM supplementation was likewise associated with decreased heat production and related heat distress mortality, and improved energetic efficiency. Belay and Teeter [25] suggested that observed response to VM supplementation was partly attributable to improved temperature homeostasis of chickens under conditions of heat stress. Consistent with this earlier work involving poultry, we likewise found that VM supplementation does in fact lower core temperatures in cattle. This is the first time that this observation has been reported for ruminants.
Trial 2. Characteristics of digestion and ruminal fermentation
Treatment effects on characteristics of digestion are shown in Table 4. There were no treatment interactions (p>0.10). Supplemental VM did not influence (p>0.10) characteristics of ruminal and total tract digestion of nutrients. This result is, in general, consistent with previous studies [2â4], although in some instances [2] VM supplementation was observed to reduce ruminal degradation of feed N.
Diet energy density did not influence (p >0.10) ruminal and total tract fiber digestion. This finding is consistent with numerous previous studies evaluating the effect of forage levels on fiber digestion in finishing diets for feedlot cattle [21,26â29]. Percentage ruminal starch digestion was greater (9%, p<0.01), and total tract starch digestion tended to be slightly greater (p = 0.07) for HED vs LED diets. This effect was not expected. Salinas-Chavira et al [21] likewise observed decreased ruminal starch digestion with increased forage level in the finishing diet. However, in numerous other studies evaluating forage levels in steam-flaked corn-based finishing diet [26â29], forage level did not affect ruminal or total tract starch digestion.
As expected, due to the greater proportion of steam-flaked corn, the HED diet resulted in greater ruminal (p = 0.02), post-ruminal (p<0.01) and total tract digestion (p<0.01) of OM, and dietary digestible energy (p<0.01). However, in spite of increased ruminal OM digestion, both ruminal microbial efficiency (p<0.01) and net flow of microbial N to the small intestine (p = 0.03) were depressed with HED vs LED diets. The observed reduction in microbial efficiency was expected. Based on NRC [5] the predicted microbial efficiency for the LED and HED diets are 25.2 and 22.2, respectively, in good agreement with observed (Table 4).
Ruminal degradation of feed N was greater (p = 0.02) for HED vs LED diets. The observed degradation of feed N for the HED diet was in good agreement (99%) with expected (NRC [5], Table 1). In contrast, observed ruminal degradation of feed N with the LED diet was 88% of expected. The basis for this lower-than-expected feed crude protein degradation is not certain. But in light of the decreased ruminal starch digestion with that diet, we speculate that the greater forage level might have produced an increased rate of passage of distillers dried grain (DDG) and associated corn. The combined effect of decreased percentage ruminal degradation of feed N and increased microbial efficiency was a tendency for increased (6%, p = 0.10) ruminal N efficiency (N entering the small intestine/N intake).
Treatment effects on ruminal pH, ruminal VFA molar pro portions and estimated methane production are shown in Table 5. Dietary energy density did not affect (p>0.10) ruminal pH, VFA molar proportions or estimated ruminal methane production. Forage levels were 17% and 10% for the LED and HED diets, respectively (Table 1). In previous work conducted at this center, increases in forage levels of steam-flaked corn-based growing-finishing diets from 10% to 20% [27] increased ruminal pH, acetate:propionate molar ratio and methane. However, when forage levels were varied within a narrower range (between 8% and 16%), forage-level effect on ruminal pH and VFA molar proportions was non-appreciable [28,29]. Additionally, the DDG content of the LED diet was 5 percentage units greater than that of the HED diet. Carrasco et al [30] observed that notwithstanding its negligible starch content, as DDG replaced steam-flaked corn in the finishing diet ruminal pH did not increase. Supplemental VM did not affect (p>0.10) ruminal pH, VFA molar proportions or estimated methane production. Likewise, Montano et al [2] and Salinas-Chavira et al [4] did not observe and effect of VM on ruminal VFA in steers fed steam-flaked corn-based finishing diets.
CONCLUSION
The magnitude for enhancement in the NE value of growing-finishing diets due to The VM supplementation may be dependent on the energy density of the growing-finishing diet. Virginiamycin-induced enhancement in dietary NE appears to be largely associated with increased efficiency of energy utilization, as effects of virginiamycin supplementation on characteristics of ruminal and total tract digestion were not appreciable. Under conditions of high ambient temperature virginiamycin supplementation may reduce core body temperature.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







