



INTRODUCTION
Intestinal epithelium not only allows selective permeability as a filter for absorption of nutrients, but also is regarded as the first line of defense to foreign antigens, such as pathogens and toxins coming from intestinal lumen (Groschwitz and Hogan, 2009; Suzuki, 2013). Tight junctions (TJs) are the apical-most adhesive multiple protein complexes that link the apical side of intestinal epithelial cells (IECs) preventing the diffusion of antigens freely across the epithelium. However, when IECs are damaged and disrupted by external stimuli including toxins, TJ alternations increased permeability and decreased integrity of the barrier (Ohland and Macnaughton, 2010). In animals, including pigs, the barrier function of intestine is crucial at the time of weaning, since the barrier can be damaged easily by solid-phase diet or infectious agents (Wijtten et al., 2011; Hu et al., 2013). Therefore, improvement of TJ formation is considered as one of the most important steps of forming an intestinal mucosal barrier to prevent paracellular diffusion of microorganisms, toxins, and other harmful antigens across the epithelium.
Deoxynivalenol (DON) is a trichothecene produced by the plant pathogens Fusarium graminearum and F. culmorum, which is prevalent in crops like wheat, oats or barley (Pinton et al., 2009). Pigs are a susceptible animal species to this toxin and ingestion of feed contaminated with DON leads to reduction in growth and immune-modulating properties and thus to economical loss (Pestka, 2008). High concentrations of DON on the enterocyte barrier results in the cell death and loss of the integrity of the epithelial barrier (Diesing et al., 2011). In the gastrointestinal tract, DON contacts the epithelial surface and is rapidly absorbed (Prelusky et al., 1988). Also, DON interferes with the expression of TJ proteins in porcine intestinal cells (Diesing et al., 2011) and binds to ribosomes inhibiting translation (Ehrlich and Daigle, 1987), suggesting the need for the development of a strategy to protect the porcine intestine from the toxin.
Probiotics are commonly used in pig feeding to stabilize the gut microflora as a preventive method. B. subtilis, gram-positive bacteria, is a soil and water saprophyte and endospore-producer that is considered as a beneficial probiotic bacteria (Hong et al., 2005). B. subtilis is known to induce the secretion of IL-8 from porcine intestinal epithelial cells, IPEC-J2 (Aperce et al., 2010). Furthermore, TLR2 recognizes various bacterial, fungal, viral, and certain endogenous substances that play an important role in the expression of TJ proteins of IEC (Kuo et al., 2013) and protection of IEC barrier function (Abreu, 2010) for intestinal homeostasis in humans and mice. However, the impact of B. subtilis together with the TLR signaling pathway on porcine intestinal barrier function is in need of further investigation.
In the present study, we hypothesized that B. subtilis may influence the expression of TJ proteins in porcine epithelial cells that could attribute to a protective effect against DON-induced damage.
MATERIALS AND METHODS
Cell culture
Non-transformed IPEC-J2 cell line (DSMZ, Braunschweig, Germany) was cultured in DulbeccoŌĆÖs modified Eagle medium (DMEM:HamŌĆÖs F-12 [1:1]) supplemented with 5% fetal bovine serum (FBS), 1% insulin-transferrin-selenium-X (ITS-X) and antibiotics (all from Invitrogen, Grand Island, USA) in an incubator with atmosphere of 5% CO2 at 37┬░C. To examine the effect of B. subtilis in the process of barrier formation, 5├Ś105 cells were cultured on 6-well plate for 3 days until reaching the confluence. For the electrical resistance experiment, the cells were grown on 0.4-╬╝m polyester membrane trans-well (Corning, New York, USA) at a density of 5├Ś104 cells/well for 3 days.
Bacteria
Probiotic strain, B. subtilis HB3 (ATCC, Manassas, USA) was cultured on trypticase soy agar (BD Biosciences, San Jose, USA) plate at 37┬░C for 12 h, and then one colony on the plate was picked and diluted in tryptic soy broth and incubated at 37┬░C for 15 h. The bacteria were collected by centrifugation at 4,000 g for 15 min, washed twice with phosphate-buffered saline (PBS), resuspended, and then heat-inactivated at 120┬░C for 30 min.
DON treatment
DON (Sigma, Missouri, USA) was diluted in absolute ethanol to prepare 2 ╬╝g/mL solution and treated to the apical side of cell monolayer for 48 h. To evaluate the effect of whole bacteria and TLR2 agonist on the barrier function, IPEC-J2 cells were pretreated with 1├Ś109 cfu/mL of heat-inactivated B. subtilis (inactivation confirmed after the plating, data not shown), 10 ╬╝g/mL of lipoteichoic acid from B. subtilis (LTA-BS; Invivogen, San Diego, USA) or media for 1 h before the DON treatment.
Electrical resistance measurements
IPEC-J2 cells were grown on 0.4-╬╝m polyester membrane trans-well (Corning) until reaching the confluence of monolayer and treated with B. subtilis or LTA-BS for 1 h. And then, during barrier formation, transepithelial electrical resistance (TEER) was measured to examine the changes in integrity between the apical and basolateral side of the cells using an epithelial voltohm meter (EVOM; World Precision Instruments, Sarasota, USA). The resistance of media as a control was subtracted from each experimental value.
Western blot analysis
IPEC-J2 cells, pretreated with LTA-BS, B. subtilis or media for 1 h, were incubated with DON for 48 h. Then, the cells were washed with PBS and lysed in lysis buffer (20 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100), followed by quantitation of protein using Micro BCA kit (Thermo, Rockford, USA). The same amount of protein extract was loaded in 10% Tris-glycine polyacrylamide gels and electrophoresed. Then, the proteins were transferred onto a polyvinylidene difluoride (PVDF) microporous membrane for 2 h at 4┬░C and blocked with 5% skim milk in TBS-T (20 mM Tris HCl, 100 mM NaCl, 0.05% Tween 20) for 90 min. The blots were incubated with rabbit anti-claudin-3, -occludin and -ZO-1 antibodies (Invitrogen) or mouse anti-╬▓-actin monoclonal IgG1 antibody (Santa Cruz Biotechnology, Grand Island, USA) overnight. Subsequently, the membrane was washed and incubated with goat anti-rabbit and -mouse IgG-HRP (Santa Cruz Biotechnology) (diluted at 1:10,000) for 1 h. The target proteins were visualized by enhanced chemiluminescence (ECL) system (GE Healthcare, Waukesha, USA) followed by analysis using ChemiDoc XRS (Bio-rad, Hercules, USA). Intensity of the blotting was quantified using Multi Gauge software (Fujifilm, Tokyo, Japan).
Confocal immunofluorescence microscopy
Monolayer of IPEC-J2 cells were pretreated with B. subtilis, LTA-BS or media for 1h and treated with DON for 48h. The cells were washed with PBS and fixed with 4% paraformaldehyde at 37┬░C for 20 min, followed by blocking with 10% FBS for 1 h at 37┬░C. The cells were stained with rabbit anti-occludin or -ZO-1 antibodies for 1 h at room temperature, followed by goat anti-rabbit IgG conjugated with Alexa fluor 488 (BD Biosciences). Nuclei were stained with 4ŌĆ▓, 6-diamidino-2-phenylinodele (DAPI) (Immunobioscience, Raleigh, USA). The distribution of target proteins was analyzed by a confocal laser scanning microscope, LSM700 (Carl Zeiss, Germany).
Statistical analysis
Unpaired studentŌĆÖs t test was used to analyze significant differences between two groups. Two-way ANOVA was used to determine differences among multiple groups for western data. TEER data were analyzed using the SAS statistical package (version 9.3; SAS Institute Inc., Cary, NC, USA). The p<0.05 was considered statistically significant.
RESULTS
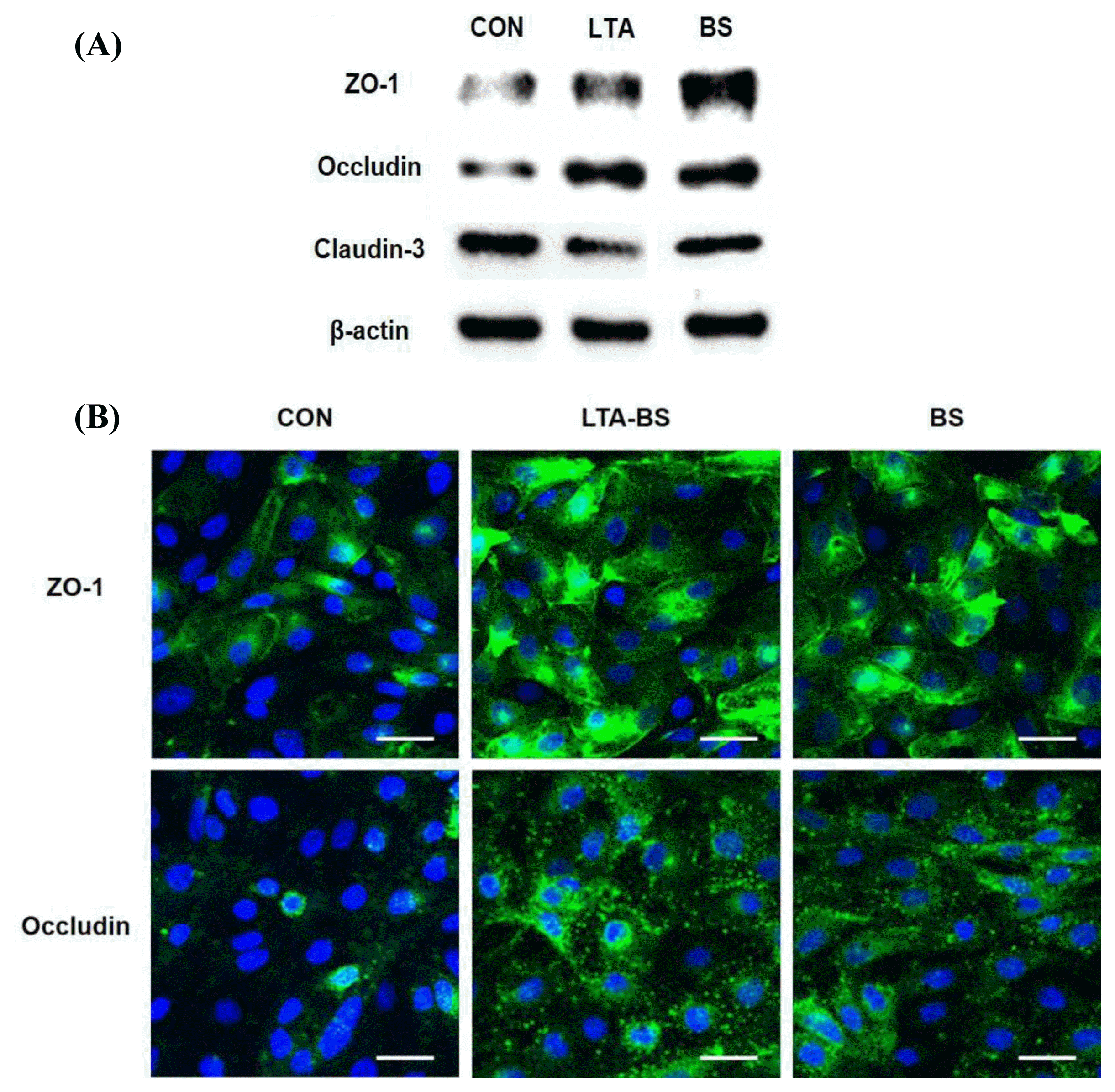
B. subtilis increases the expression of ZO-1 and occludin in IPEC-J2 cells
Previously, LTA was shown to enhance TJ protein expression in the mouse (Kuo et al., 2013). Furthermore, our study demonstrated that 24 h treatment with LTA-BS enhanced the expression of tight junction proteins, including ZO-1, occludin and claudin-3 in porcine epithelial cells, and protected against 48 h DON exposure (unpublished data). In the present study, to investigate whether the short-term treatment with LTA-BS or B. subtilis also promotes the expression of TJ proteins, we treated IPEC-J2 cells with LTA-BS or B. subtilis for 1 h and measured the protein changes of ZO-1, occludin and claudin-3. The expression of ZO-1 and occludin was up-regulated in the cells treated with LTA-BS or B. subtilis when compared to those in control, while claudin-3 expression minimally reduced (Figure 1A). Furthermore, confocal imaging confirmed that ZO-1 and occludin expression in the cells treated with LTA-BS or B. subtilis were improved compare to those of control (Figure 1B). It was noted that ZO-1 formed a clear outer-most boundary of the cells when treated with B. subtilis. Therefore, we found that short-term treatment of IPEC-J2 cells with B. subtilis induced an increase in the expression of TJ proteins, ZO-1 and occludin.
B. subtilis induces increase in ZO-1 expression against damage from DON treatment
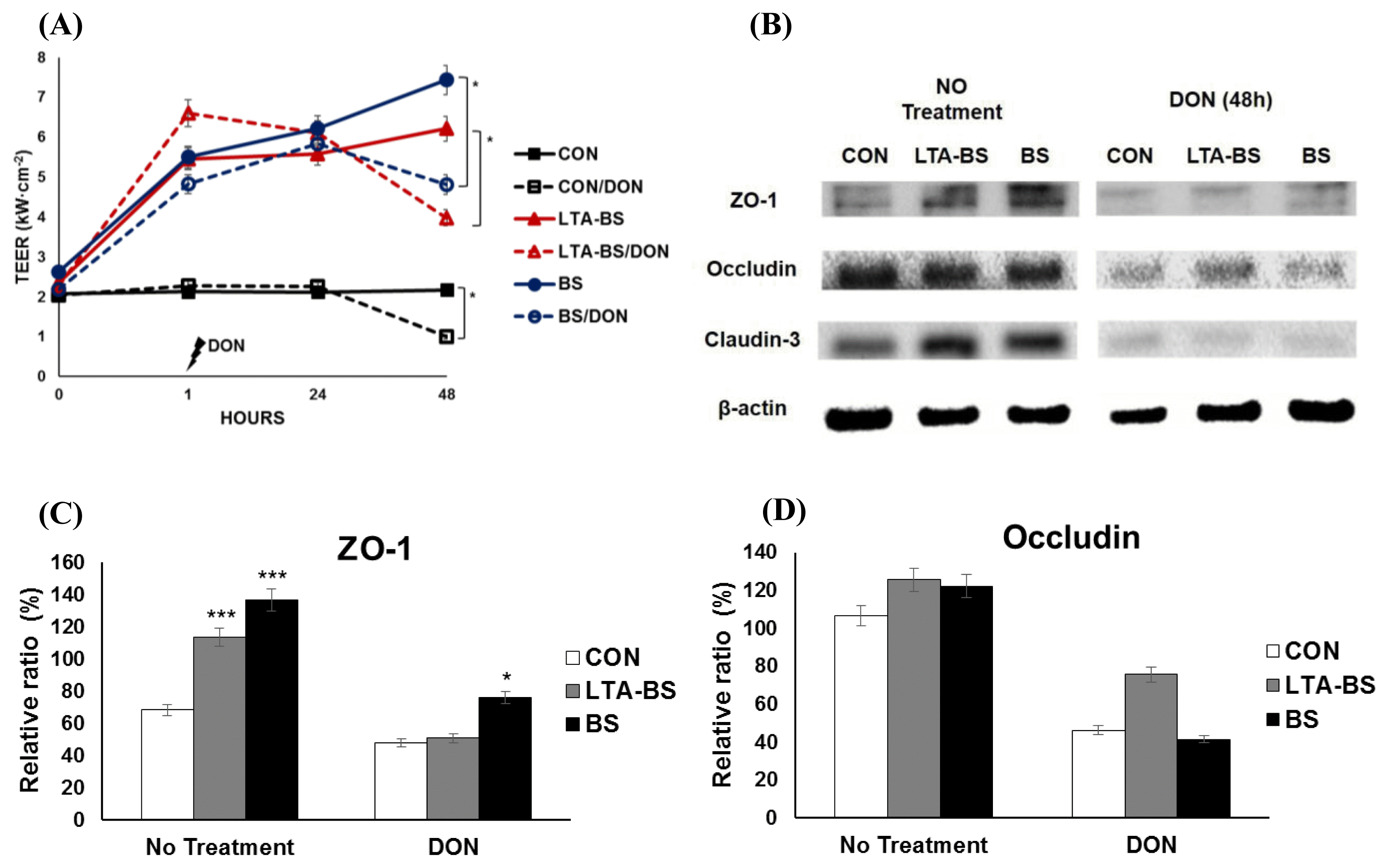
DON is known to exhibit toxic effects in pigs by increasing permeability of the intestinal epithelial cell barrier (Pinton et al., 2009). To examine the cell survival against DON treatment, cell death was examined by using flow cytometry analysis. The results showed that 2 ╬╝g/mL of DON exposure for 48 h was enough to induce the damage to the IPEC-J2 cells without inducing cell death (data not shown). To evaluate the protective effect of B. subtilis on the permeability of IPEC-J2, TEER was measured on monolayer cells and the result showed that the cells treated with LTA-BS or B. subtilis for 1 h increased TEER compared to those not treated (Figure 2A). Besides, DON treatment for 48 h led to significant drop in TEER in comparison with no treatment (Figure 2A).
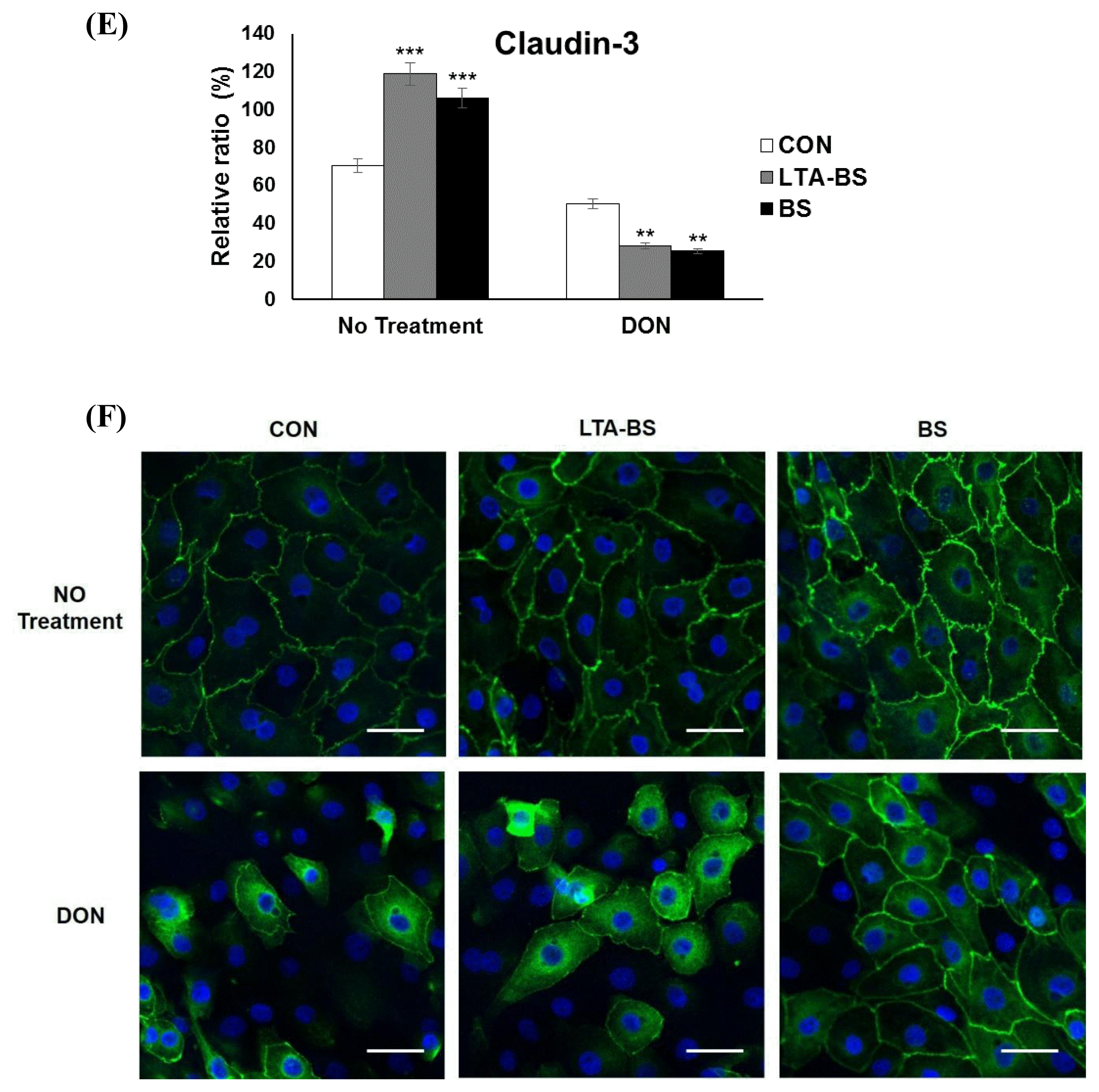
We found that B. subtilis and LTA-BS provides resistance against permeability decrease in the porcine epithelial cells treated with DON. To analyze the change of tight junction proteins, we examined the expression of ZO-1, occludin and claudin-3 in IPEC-J2 cells which were pretreated with LTA-BS or B. subtilis and exposed to DON. The result indicated that the cells, treated with LTA-BS or B. subtilis (without DON treatment), expressed more ZO-1 (Figure 2B and C) and claudin-3 (Figure 2B and E) than those of control. DON-treated cells showed decline in TJ protein expression (Figure 2B, C, D, and E). Interestingly, pretreatment with B. subtilis appeared to induce better expression of ZO-1 than other proteins, which was revealed by confocal imaging that showed ZO-1 retained outline formation of the cells. In contrast, control or LTA-BS-treated cells became round without being associated each other, and TJ protein was expressed near nucleus, not boundary edge of the cells (Figure 2F). These results coincided with the increased TEER on B. subtilis-pre-treated cells against DON compared to that of control (Figure 2A).
DISCUSSION
We proposed, in the present study, that short-term treatment with B. subtilis, as a whole bacteria, could promote the formation of TJ proteins and function of the epithelial barrier. To this end, we evaluated the effect of B. subtilis on the expression of TJ proteins on IPEC-J2 cells for 1 h, prior to 48 h DON exposure. The results showed that IPEC-J2 treated with B. subtilis recovered and maintained the expression of ZO-1 compared to LTA-BS and control. It has been proposed that enhancement of barrier formation of IPEC-J2 could decrease the permeability against the damage by toxins including DON.
DON is known to specifically target dividing cells like IEC and immune cells (Pestka et al., 2004). DON binds to RNA peptidyltransferase site and inhibits protein synthesis (De Walle et al., 2010). It induces expression changes of transcription factors by binding to the ribosomes and activating MAPK pathway (Zhou et al., 2003). Through stimulation of p44/42 extracellular signal-regulated kinase and C-Jun-N-terminal kinase, DON induces alteration of TJ proteins and intestinal barrier dysfunction in mice (Kolf-Clauw et al., 2009; Diesing et al., 2011). In the case of pigs, DON showed a negative effect on the expression of TJ proteins of IPEC and its toxicity could involve a down-regulated expression of ZO-1, occludin and claudin-3 of IPEC-J2 (Diesing et al., 2011). However, the mechanism of such event is not yet fully understood.
TJ consists of transmembrane proteins including claudins, occludin and junction-associated molecules. Additionally, there are intracellular proteins called ZO proteins which are in conjunction with cytoskeletal actin and actomyosin (Yu et al., 2012). We found that DON impaired the expression of TJ protein and B. subtilis protected against a decline in ZO-1 expression. ZO-1 interacts with claudins, occluding, and actin (Turner, 2009), and plays an important role in structural organization of tight junctions (Stevenson et al., 1986). ZO-1 deficient cells showed characteristic of delayed assembly with other TJ proteins (Guan et al., 2011). Although occludin is a crucial constituent of TJ proteins, the formation of TJ is not solely dependent on occludin as shown by using occludin-deficient mice (Suzuki, 2013). It was further confirmed that claudin is not always necessary for the function of TJ. It has been suggested that activation of protein kinase C via TLR2 pathway improved the tightening of TJ associated with ZO-1 in the intestinal epithelial barrier, but not occludin, claudin, or actin cytoskeleton (Cario et al., 2004). Therefore, in our study, B. subtilis-enhanced expression of ZO-1 is coincident with up-regulated formation of epithelial barriers as indicated by the resistance of the TJ barrier against DON damage.
It has been demonstrated that TLR2 stimulation exerted TJ-protective effect via MyD88-dependent PI3K/Akt-pathway (Cario, 2008). TLR2 agonist enhanced the expression and protective function of TJ proteins in human keratinocytes (Kuo et al., 2013), however, LPS (i.e., TLR4 agonist) stimulation does not activate PI3K/Akt signaling and regulation of membrane integrity (Cario et al., 2007). In addition, TLR2-induced PI3K/Akt (Cario, 2008) and p38 MAPK (Ueno et al., 2011) pathways are related to NF-╬║B translocation that regulates the function of TJ barrier in mice and humans. We found that IPEC-J2 also had enhanced expression of TJ proteins through PI3K/Akt signal by TLR2 ligands, including LTA-BS, peptidoglycan from B. subtilis (PGN-BS) (unpublished data). Thus, we proposed that B. subtilis, which has TLR2 ligands, would promote TJ protein expression. In the present study, we used LTA-BS as a positive control and observed that B. subtilis, similar to the effect of TLR2 agonist on IPEC, strengthened the function of the porcine intestine epithelial barrier through enhanced expression of TJ proteins.
Furthermore, Lactobacillus casei prevented the damage in Caco-2 cells through the increase of the ZO-1 expression via activation of p-Akt (Eun et al., 2011). Pretreatment with gram-positive bacteria, L. rhamnosus and L. helveticus, reduced pathogenic bacterial adherence and translocation (Zareie et al., 2006). It is probable that B. subtilis might have a potential to induce improvement of intestine barrier function. Whole bacteria are equipped with various ligands for receptors including TLRs, formylated peptide receptors (FPRs), nucleotide binding oligomerization-like receptors (NODs), IL-1╬▓-converting enzyme protease activator factor (IPAF), and C-type lectins (Turner, 2009; Ohland and Macnaughton, 2010), which could exert a diverse effect on intestinal barriers. However, aforementioned studies are all related to mice or humans, and therefore, we are in need of further investigation on the clarification of B. subtilis-induced pathways including TLR2 in epithelial cells of pigs.
In fact, B. subtilis allowed IPEC-J2 to express higher ZO-1 than did LTA-BS and to form TJ proteins along the outside of the cells in our study. It suggested that B. subtilis containing multi-innate receptors might exert a higher protective effect on TJ barrier than LTA-BS alone. Thus, B. subtilis could be an effective candidate for improvement of porcine intestinal barrier function. It is worthwhile to note that considering the expression of other TJ proteins and the degree of formation of TJ, pretreatment of the epithelial cells for 1h could be too short for fully effective protection, in spite of the fact that the expression of ZO-1 increased in porcine epithelial cells.
In conclusion, B. subtilis seemingly improved porcine epithelial barrier function by preventing overwhelmingly increased permeability and promoting TJ protein expression. In addition, B. subtilis had a protective role against DON-induced barrier impairment through the up-regulated expression of ZO-1. Collectively, the present study suggests an important implication for the role of B. subtilis on the regulation of tight junction proteins in porcine intestine epithelial barrier, and that is a potential candidate as a feed additive to protect against intestinal damage in pigs.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







