



INTRODUCTION
Hypothermia is one of the major cause of neonatal piglets’ mortality. At birth, piglet is exposed to drastic changes in the ambient temperature. Considering the physiological temperature in sow’s utero, between 38°C to 40°C, immediately after the piglets’ birth, whose minimum temperature comfort is around 34°C to 35°C (Mount, 1959; Manno et al., 2005), they come in contact with a colder environment in farrowing crates, near the comfort range for lactating sow, 18°C to 23°C (Yan and Yamamoto, 2000; Brown-Brandl et al., 2001). This event triggers the reduction of body temperature soon after birth (Tuchscherer et al., 2000; Pandorfi et al., 2005; Malmkvist et al., 2006).
This process is known as post natal hypothermia and its extension and duration is negatively correlated with the survival chances of the piglet (Tuchscherer et al., 2000). These losses can affect piglet in the ways of as low intake of colostrum, low development, higher susceptibility to disease and more cases of crushing (Pandorfi et al., 2005; Souza, 2007). Nevertheless, there are large individual differences between and within litters, in the success of newborn piglets’ recovery (Tuchscherer et al., 2000). The recovery of body temperature to normal physiological values depends on factors such as room temperature, piglet’s weight at birth, time required to start breastfeeding and the management adopted in the farm.
Currently, strains of sows intended for breeding are being genetically improved in order to become hyper prolific. Furthermore, changes inherent to management allowed the increase in number of weaned piglets per female per year, from an average 21 to 23 piglet (Mercks et al., 2000) to a level surrounding 28 to 30 weaned piglets per female per year (Antunes, 2007). Consequently, there was an increase of problems related to birth weight and nonuniform litters, contributing to the higher variability in weight between piglets (Sorensen et al., 2000; Damgaard et al., 2003).
The lower birth weight predisposes piglets to lower chances of surviving (Van Rens et al., 2005) and this effect is more pronounced in piglets with an average weight less than 1.0 kg (Quiniou et al., 2002). These piglets have lower levels of body energy supplies, greater sensitivity to cold, take longer to achieve the mammary complex and a lower ability to choose the best teat (Lay Júnior et al., 2002).
The thermoregulatory capacity is directly related to birth weight. Lighter piglets have higher body surface in relation to its weight, and they are, therefore, more likely to be affected by hypothermia (Herpin et al., 2002). Panzardi et al. (2009) found that birth weight and body temperature at 24 h post-birth are among the best predictors of survival rate during the first postnatal week.
Thereby, this study was carried out in order to verify the effects of piglets’ birth weight in individual change on surface temperature after birth, and to determine its relationship with the time required for colostrum ingestion.
MATERIAL AND METHODS
Local, experimental design and animal management
The experiment was conducted in a full cycle commercial pig farm, located in Dourados, MS. The city altitude is 430 m, latitude 22° 13′ S and longitude 54° 48′ W. The climate is tropical with rainy summer and dry winter.
Litters from four sows, crossbred Landrace×Large White, were used in the research, all of third farrow. The females were distributed in individual farrowing cells with a total area of about 4.0 m2, provided with protection against crush and a creep was provided with incandescent lamp as a heat source for the piglets.
The production system works with forced synchronization of birth, with an average of six to seven calving per week. Females were induced through the injection of 0.12 mg cloprostenol via SMV, with 24 h prior to expected date of parturition.
After birth, the piglets were instantly dry with paper towel and received numbers with ink marker. After the rupture of umbilical cord, that occurred naturally, without assistance of the workers, the piglets were weighed and sent for first colostrum ingestion, which was recorded, for each animal. This variable was defined as the first time the piglet breastfed continuously in the same teat for at least one minute.
From the birth weight, the piglets were divided into a completely randomized design with three treatments, which consisted in the ranges of birth weight: T1 - less than 1.00 kg, T2 − 1.00 to 1.39 kg, and T3 - higher than or equal to 1.40 kg.
Infrared thermography and evaluation of piglets’ surface temperature
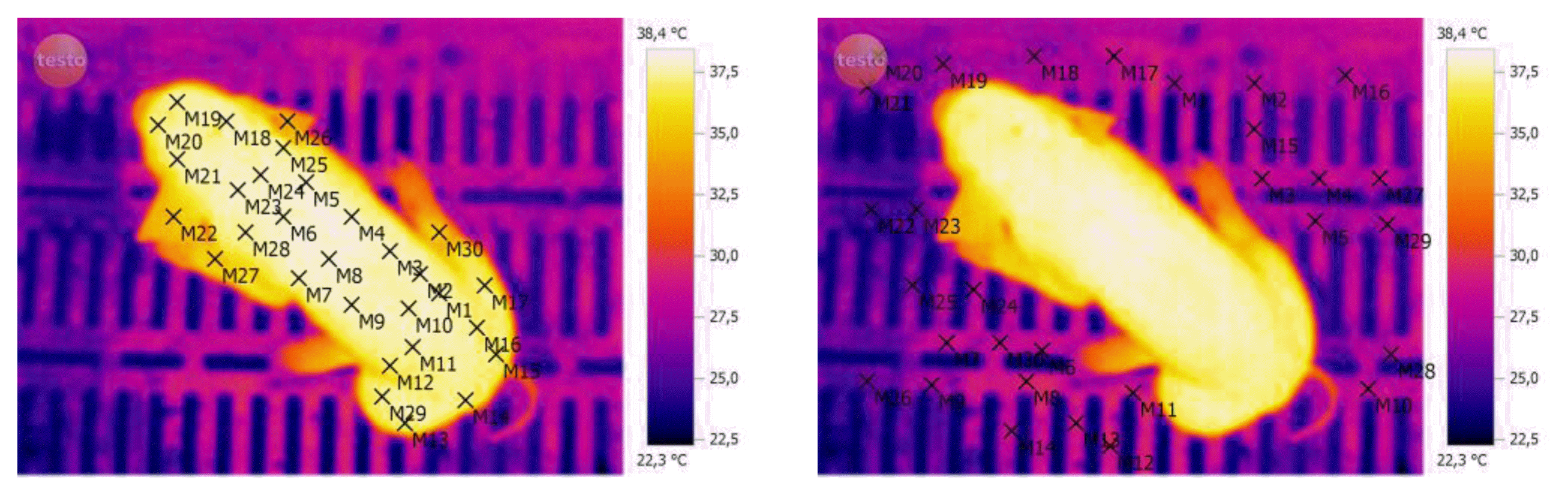
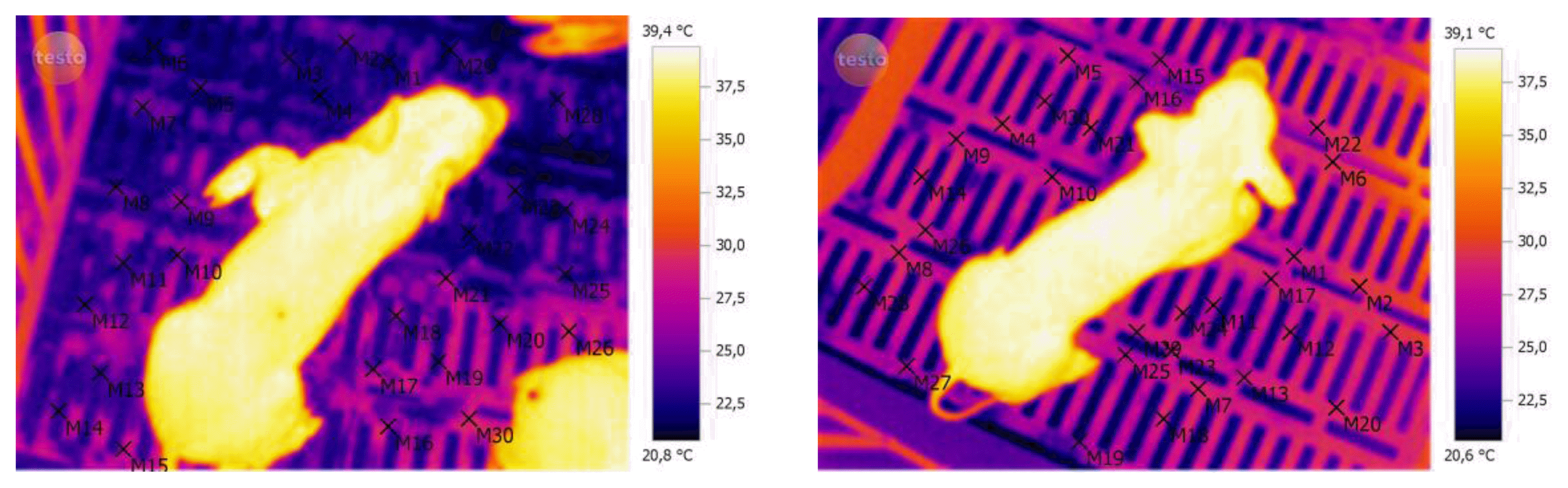
Infrared images of all piglets were registered at birth (after performing the cleaning procedure) at 15, 30, 45, 60, 120, and 180 min after birth. The thermal images recording was performed using the Termovisor Testo 876-1 Kit Professional equipment, with accuracy of ±0.1°C, and the images were processed using the Testo IRSoft software Version 3.1 SP2. The emissivity coefficient of 0.96 was used for all body surface of the animal. The average and standard deviation values of temperature of the body surface were calculated using the 30-point temperature selected to represent entire body surface of the animal, or by selecting areas to construct a histogram temperature (Figure 1). The surface temperature of farrowing crates floor was also recorded, using the same procedure and an emissivity coefficient of 0.94 for plastic.
Environment parameters
The temperature and relative air humidity, inside and outside the farrowing crate were recorded every 30 min, during the experimental period, using a digital thermo-hygrometer.
Using the recorded values of air temperature and relative humidity, the dry bulb temperature (dbt) and wet bulb (wbt) were calculated via Psicrom program (Roriz, 2003) and after that, the Temperature and Humidity indexes (THI) were determinate by using equation 1 described by Thom (1958):
Statistical analysis
Variance analysis was applied to the surface temperature data and time to first feeding, using initially the ranges of birth weight and thereafter sows as treatments. The averages were compared by Tukey test at 5% significance, using SAS 9.2 (SAS, 2001). Spearman correlation analyzes between piglet’s weights at birth, the surface temperature over time after birth and time to first feeding were performed.
RESULTS AND DISCUSSION
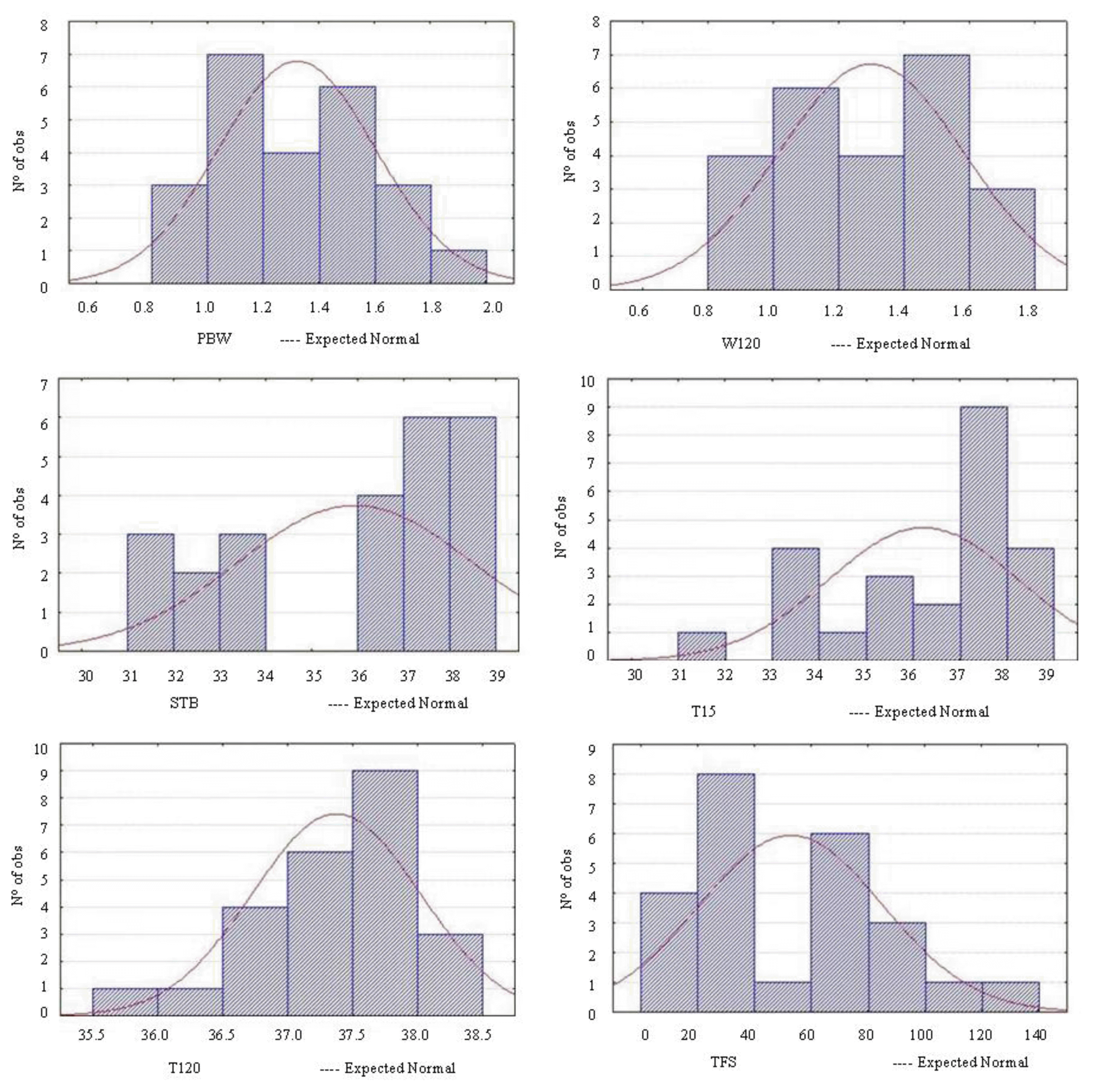
The studied variables showed similar data and slight variations in averages combined with low coefficient of variation values (CV%), indicating sample precision, except for the time for first feeding (TFS), which presented a considerable variation, and thus showed a higher data dispersion (Table 1).
Observing the piglet weight after two hours old (P120), it was clear that there was a slight change compared to birth weight (Figure 2). There was a decrease in ST after the first 15 min of life, increasing again two hours after. The largest drops in temperature occurred between fifteen and sixty minutes after the birth of piglets (between 2.15°C to 5.61°C, average 3.88°C), normal values (1.7°C to 6.7°C with an average 2.2°C), compared to reference ranges according to Mendonça (2010).
Environment
The mean temperature inside the farrowing crate from 10:00 am to 6:00 pm ranged from 26.3°C to 32.5°C and the relative humidity ranged from 34.3% to 47%, reaching highest temperature values and lowest humidity between 3:00 pm and 4:00 pm.
The ideal environment to be provided for pigs varies according age and physiological condition. In the specific case of sows farrowing, there are two animal categories with different comfort temperature ranges: the lactating sow, whose comfort range is 18°C to 23°C (Yan and Yamamoto, 2000; Brown-Brandl et al., 2001); and the piglet, which comfort range is 30°C to 32°C (Lima et al., 2011). The increase in internal temperature of farrowing rooms in order to attend the thermal demand of piglets very often disregards the temperature range of comfort for the sow, which can cause heat stress in the female, affecting milk production (Lima et al., 2011), and interfering with the natural process of birth due to heat stress and suppression of secretion of oxytocin. However, this fact was not observed in this study, and no sows in this trial showed problems with dystocia during the birth process, which lasted around 2 to 3 h.
When considering the recommended values of air temperature and relative humidity for piglets in the first week of life, the THI within the thermal comfort ranges from 83.5 to 87.6, with critical values of THI less than 66.4, and higher than 92.1. Thereby, calculated values (75.63 to 82.51) were found during most of the time, closer to the thermal comfort zone than to the critical range; however, they were below the ideal ones.
Birth weight effects
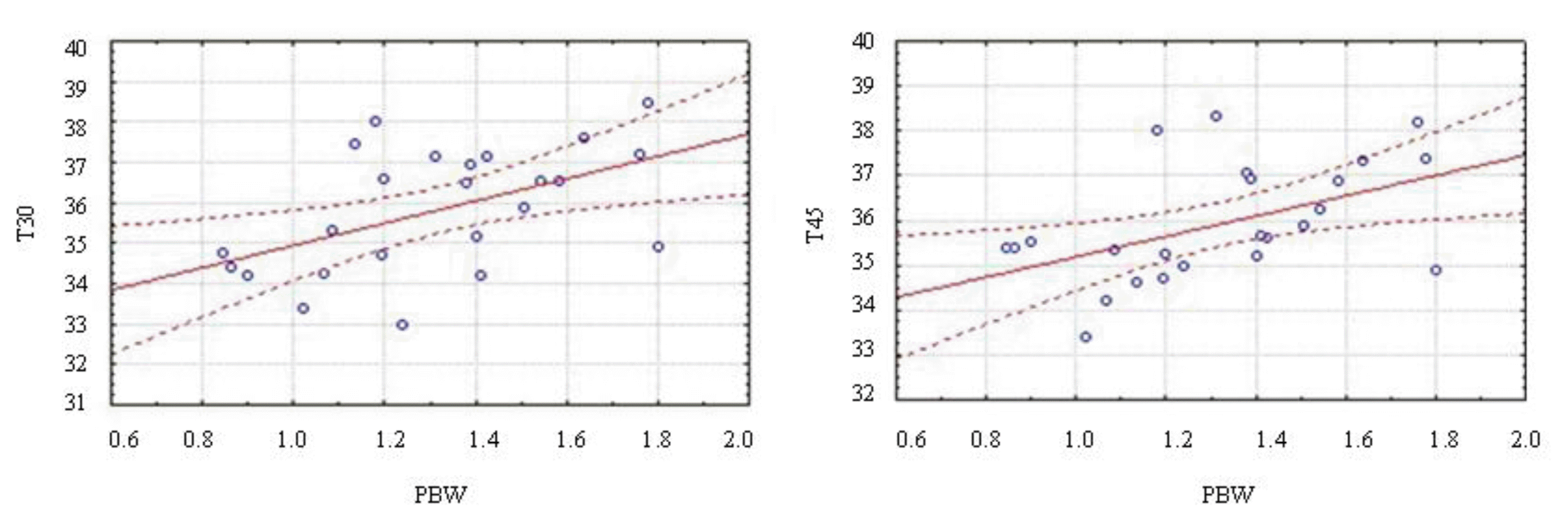
Positive correlations were observed between piglet weight at birth (PBW) and their surface temperatures at 30 and 45 min after birth, indicating that the heavier the piglet’s birth weight, the higher surface temperature during this time interval, and then, the lower drop in surface temperature due to hypothermia postnatal (Table 2, Figure 3). Therefore, since the fall of body temperature in the first 60 minutes after birth is observed, the first hours of life in the piglets are critical times for the animal, especially for those with low birth weight.
Hypothermia occurs naturally after birth, in the majority newborn mammals, but both the extent of initial drop in body temperature and time required for subsequent recovery are highly variable. Newborn piglets are inadequately shielded, lacking of brown adipose tissue and depend on almost exclusively on shivering to thermogenesis for cold (Herpin et al., 2002).
At birth, piglets usually have sudden reduction in body temperature and the recovery of normal temperature of 39°C is achieved within the first 24 to 48 h after birth, being directly dependent on the temperature of their body weight and the moment he begins to suckle (Quesnel et al., 2012). However, excessive hypothermia due to adverse environmental conditions, low birth weight or reduced vitality can reduce the vigor and colostrum intake and lead, ultimately, death of the animal (Souza, 2007; Panzardi et al., 2013). By exposing newborn piglets to hypothermia, Lossec et al. (1998) observed that the body temperature and skin showed, rigorously, the same behavior, suggesting that even though the concentration of circulating catecholamine have increased during cooling of the body piglets were unable to reduce heat losses effectively through peripheral vasoconstriction.
When assessing connections between different physiological and behavioral measures (birth weight, birth hypoxia, latency between the first udder contact and first feeding) and connections with piglets’ ability to overcome hypothermia early postnatal, Jensen et al. (2011) concluded that birth weight is the most important factor in successful recovery from hypothermia postnatal. The correlation between birth weight and thermoregulation capacity of piglets in challenging situations by cold was also described in other researches (Herpin et al., 2004; Gondret et al., 2005; Baxter et al., 2008). Small piglets have higher surface area relative to the volume of the body in comparison with the large piglets, and these animals are, consequently, more prone to heat loss in a cold environment. Thus, small piglets are especially at increased risk since the heat loss per weight unit is inversely related to the size of the body. The low capability to retain heat is reflected by fact that each 1°C reduced below the minimum critical temperature is associated with increased from 1.46 kJ kg−0.75h−1 heat production, a value three times higher than in 35 kg pig (Herpin and Le Dividich, 1995). Differences in weight related to physiological maturity may be another possible explanation for the relationship between the success of thermoregulation and birth weight (Jensen et al., 2011).
Relating to birth weight and efficiency of thermoregulation, Jensen et al. (2011) and Herpin et al. (2004) suggested that there was a minimum birth weight close to 1,100 g, and below that modern European breeds piglets have thermogenic capacity linearly impaired.
The average surface temperature of the floor in farrowing crates ranged from 24°C to 28°C. (Figure 4). Considering that much of the heat loss by piglets at that moment occurred by conduction, we realized the importance of management practices to minimize heat exchange between the neonates and the floor, such as the use of bedding.
Birth weight was negatively correlated with the time to first suckle and spent (TFS), suggesting that piglets with higher birth weight ingest colostrum faster than piglets with lower weight (Table 2), which was similar to that observed by Jensen et al. (2011).
The body energy stored at birth is present in the form of glycogen and fat. However, the available energy from glycogen and fat derivatives for newborn piglets is extremely low. Carbohydrates are the first and predominantly utilized by newborn pig and its use is significantly accelerated during exposure to cold (Orozco-Gregorio, 2007) leading to the rapid depletion of glycogen (Pastorelli et al., 2009). The amount of body fat is also extremely low at birth, and most of them are not available for oxidation. Colostrum consumption provides to piglets large amounts of fat (30% to 40% of total energy of colostrum) and, therefore, substantial changes thereby providing energy substrates for thermoregulation. Once piglets with low birth weight possess even smaller amounts of energy reserves and given their greater difficulty on ingest considerable amounts of colostrum special attention should be given to minimize these losses by hypothermia.
High correlation was found between birth weight and weight after two hours of life (P120), which was already expected since there was low variation between the weights of piglets in this period. Thus, the time to first suckle (TFS) also showed high negative correlation with the P120.
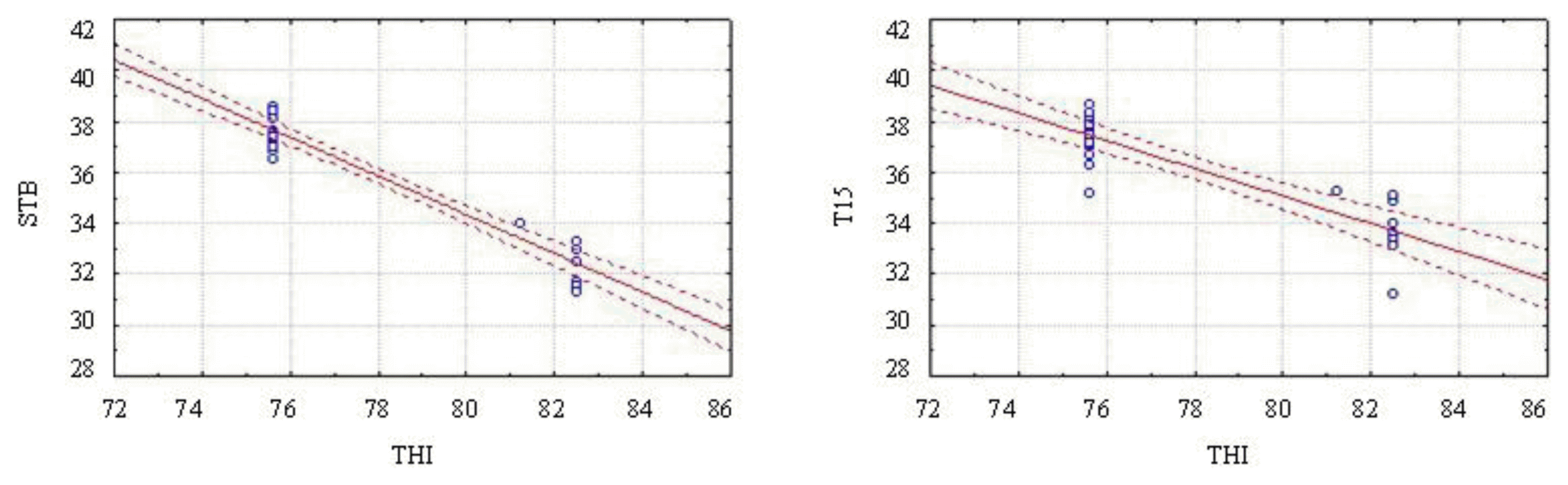
THI showed high negative correlations (−0.824 and −0.815) (p<0.01) with the surface temperature at birth (TNS) and at 15 min after birth (T15), indicating that the higher the THI the lower the surface temperatures of piglets in the first moments after birth (Table 2, Figure 5). This result shows that the ambient has a direct influence on piglet body temperature, since heat stressed sows may show physiological changes in an attempt to maintain thermal homeostasis, reflected during delivery, directly in the fetuses.
The piglets’ temperature at birth was positively correlated with temperature at 15 min post-natal influencing consequently the temperatures in the interval of 45 to 120 min. Between 60 and 120 min after birth, there was an increase in the temperature of the animal, explaining the high positive correlation (0.614) (p<0.01) between these variables. This rapid recovery attempt in body temperature between one and two hours after birth may reflect good thermal conditions of the accommodation on the farrowing moment.
Individual effect of the sow
With the intention to evaluate individual effects of mothers on the surface temperature behavior of piglets and time to first feeding, females were considered as treatments. Piglets’ birth weight showed no significant difference (p> 0.05). However, the piglets’ surface temperatures from birth to 60 min were significantly different (p<0.05) for the different piglets from the sows, and the same equaled since 120 min after birth (Table 3).
A total of 25.71% of piglets showed surface temperature below 35°C at birth, 20% between 35°C and 37°C and 54.29% higher than 37°C, emphasizing that all those whose skin temperature at birth was below 35°C were from the same sow, which would characterize an individual effect of the mother.
Piglets born from the sow 4 showed the lowest surface temperature at birth (p<0.05), and hence lower temperatures at 15, 30, and 45 min post-natal, equaling to others after this period. In general, the piglets from other sows showed similar in the first two hours of life. As all the animals were kept and born in the same environment, there was no significant effect of birth weight of piglets between sows, it can be inferred the existence of genetic factors inherent to mother, which could affect piglets’ thermoregulatory mechanisms (Table 4).
Jensen et al. (2011) also observed a strong effect of sow on piglets’ rectal temperature at birth when studying factors that influence thermoregulation in neonatal pigs. However, this effect was no longer present at two hours after birth, similar to the results obtained in this study, in which the rectal temperature two hours after birth was not related to rectal temperature at birth, or with sow.
The cold stress at birth reduces the piglet strength, thereby reducing the amount of colostrum intake, and, therefore, nutrients which serve as an energy source for thermal regulation and the pig become lethargic and more likely to be crushed by the sow. This result indicates that the maintenance of homoeothermic and practice of early nutrition are of great importance for the survival of piglets. The permanent assistance at birth is a recommended practice and as the piglets are born, they must be clean and dry to reduce the loss of body heat, and avoid them to spend their few energy reserves in an attempt to maintain body temperature. Special attention should be offered to newborns with low birth weight, as they are more prone to hypothermia postnatal.
Considering that during farrowing, about 70% of piglet mortality occurs in the first week of life (Ferreira et al., 2007) and the caused are mostly by non-infectious problems such as crushing and starvation, which can be caused by exposure to cold. Thus, appropriate management practices can significantly reduce losses, and set in improvements in the profitability of the production system.
In practical terms, an additional source of heat must be offered to piglets in farrowing cages. However, they tend to stay close to their mothers and ignore this source of heat on the first day of life. Therefore, measures should be adopted to encourage the retention of piglets inside the creep in the periods in which they are not breastfed.
IMPLICATIONS
The piglet weight at birth interfere significantly in the behavior of its postnatal surface temperature and consequently in the time it takes to ingest colostrum. Heavier piglets at birth have lower temperature drop and the first intake of colostrum occurs more rapidly. Piglets with low birth weight require special attention to avoid losses due to hypothermia.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







