



INTRODUCTION
An accurate supply of energy and nutrients is essential for optimizing pig production. Therefore, accurate information about both energy and nutrient requirements of pigs and the nutritive values in feed ingredients and diets are required. One of the major objectives of pig nutrition research is to match energy and nutrient requirements for pigs with the amounts of dietary energy and nutrients in a cost-effective way. However, the recent increase in feed costs and growing concerns about environmental pollution as a result of livestock production challenges swine nutritionists to choose feed ingredients and evaluation systems for energy and nutrients in feeds that maximizes the utilization of energy in diets fed to pigs.
Energy is not a nutrient, but is required for all biological processes in pigs. Thus, an adequate supply of energy in addition to the supply of nutrients is a prerequisite for optimal pig production. The costs of ingredients supplying energy for pigs contribute the largest portion of total feed costs, and therefore, accurate estimation of both energy requirements for pigs and feed energy values may reduce the costs of pig production (Noblet et al., 1994a). Another important aspect of energy concentration in feed ingredients is that the amount of energy in diets influence voluntary feed intake of pigs because pigs have a tendency to consume feed until their energy requirements are fulfilled (Nyachoti et al., 2004). Therefore, feed energy values have been used for prediction of voluntary feed intake and subsequently for determination of needs for essential nutrients in diets. As a consequence, diets that have an appropriate ratio of energy to essential nutrients (e.g., amino acids) need to be formulated to ensure maximum profitability in pig production. During recent decades, much new knowledge has been generated on energy utilization by pigs and efforts have been directed towards developing appropriate energy evaluation systems for diets and feed ingredients. The objective of this contribution is to review current knowledge on energy utilization and energy evaluation systems in feeds for growing pigs.
ENERGY UTILIZATION
Energy is generated from the oxidation of organic compounds (i.e., carbohydrates, lipids, and proteins) in feed ingredients. Whereas nutrients provide building blocks for the animals’ body, energy is utilized for maintenance and production (Rijnen et al., 2004). In animal nutrition, therefore, energy represents an overall quality that is associated with nutrients in feed ingredients (Moehn et al., 2005). The potential energy in feeds can be stored in chemical components of the body or can be utilized for biological functions as ATP (Emmans, 1999). Energy metabolism is divided into 2 categories. Catabolism produces energy by oxidation of fuel molecules in exergonic reactions, whereas anabolism synthesizes body components in endergonic reactions by capturing the energy released from the exergonic reaction (Mayes, 2000). All processes in these exergonic and endergonic reactions must be coupled and subsequently, the overall net change in energy must be exergonic (Mayes, 2000). It is estimated that oxidation of 1 mole of long chain fatty acids, glucose, and amino acids generates approximately 112 to146 moles, 38 moles, and 6 to 42 moles of ATP, respectively (Blaxter, 1989). The energy produced from nutrient oxidation, however, is captured partially in the form of ATP and some of the energy is inevitably lost as heat during metabolism. In addition, the physiological energy that is carried by ATP is not completely utilized in the body and is also dissipated as heat when ATP is used for various biological functions.
When pigs consume feeds, dietary energy is either absorbed or excreted in feces, urine, or heat. Dietary energy absorbed by pigs is then utilized for maintenance or retention of protein or lipids (van Milgen and Noblet, 2003). It has been assumed that energy in pigs is first prioritized for maintenance and additional energy intake in excess of the energy requirement for maintenance is retained as protein or lipids in the body (Lizardo et al., 2002). The energy requirement for maintenance accounts for approximately one third of total dietary energy utilization and the remaining two thirds of dietary energy is stored as proteins or lipids in growing pigs (Black and de Lange, 1995; NRC, 1998). However, the ratio of energy utilization for maintenance or retention also depends on the growth stage of pigs, genetic background of the pigs, thermal environment, and nutritional composition of the diet.
Energy utilization for maintenance
Energy utilization for maintenance is related to basic physiological functions such as blood flows, respiration, muscle tone, ion balance, immune responses, tissue turnover, and the control of homeostasis in the body as well as physical activity, and ingestion and digestion of feeds, (Baldwin, 1995; Black and de Lange, 1995). The energy requirement for maintenance in pigs has typically been calculated as the ME requirement for maintenance (MEm) or the NE requirement for maintenance (NEm).
In general, MEm and NEm are expressed proportionally to metabolic BW as an exponential function (aBWb, NRC, 1998). The reason for adopting this expression is the proportionality between fasting heat production (FHP) and metabolic BW of animals (Kleiber, 1975). In theory, if an appropriate exponent is chosen, the proportionality is maintained between BW raised to the exponent and the energy requirement for maintenance (Chwalibog, 1991). The metabolic BW has been expressed as BW0.75 for most mammals because of a linear association between FHP and BW0.75 in mature and post-absorptive animals among different species (Kleiber, 1975). The exponent of 0.75, however, has been questioned because of inaccuracy when applied to FHP within a species or for rapidly growing animals (Thonney et al., 1976; Baldwin and Bywater, 1984). For example, it is reported that the MEm decreased with increasing BW for growing pigs when 0.75 was used as the exponent (Chwalibog, 1991). This result is likely caused by changes in body composition and in the relative size of visceral organs to total body size during the growing period (Tess et al., 1984; Noblet et al., 1991). It has been suggested, therefore, that an exponent of 0.60 is more accurate than 0.75 for growing pigs to predict the energy requirement for maintenance (Brown, 1982; Noblet et al., 1994a; van Milgen et al., 1998), and the exponent of 0.60 is currently adopted in the expression of metabolic BW for growing pigs (Noblet et al., 1994a; NRC, 2012). For sows, however, the exponent of 0.75 is still used for expressing the energy requirement for maintenance (NRC, 2012) because it has not been conclusively demonstrated that a different exponent more accurately describe maintenance energy of mature animals. It has also not been established, which exponent most accurately describe maintenance energy across various stages of production and different production rates of pigs (Chwalibog, 1991; Noblet et al., 1994b; Noblet et al., 1999).
Estimation of the energy requirement for maintenance
The energy requirement for maintenance has been determined with fasting pigs or pigs that are fed a restricted amount of energy, which leads to zero energy retention in the body (Chwalibog, 1991). However, those methods have been criticized because of differences in energy metabolism between fasting or restrictedly-fed pigs and pigs fed on an ad libitum basis (Baldwin, 1995). A more practical approach may be to determine the energy requirement for maintenance as a conceptual constant based on linear regression analysis (Verstegen, 2001; van Milgen et al., 2008). In this approach, the energy requirement for maintenance is estimated by regressing the energy retention (or total heat production) obtained for animals consuming graded levels of energy on their corresponding energy intake. The x-intercept of this regression line represents MEm and the y-intercept represents extrapolated FHP (van Milgen and Noblet, 2003; Figure 1). The extrapolated FHP is also called the operational energy requirement for maintenance (Kil, 2008; Ayoade et al., 2012).
The MEm can also be estimated from the difference between total energy intake and energy used for protein and lipid retention if the efficiencies of ME for protein or lipid synthesis are known (Birkett and de Lange, 2001). Estimates of MEm for pigs vary from 92 to 160 kcal/ BW0.75 and the average estimate for MEm is 106 kcal/ BW0.75 (NRC, 1998). When 0.60 is used as the exponent for metabolic BW, estimates for MEm range from 186 to 250 kcal/kg0.6 (Just et al., 1983; Noblet et al., 1991, 1994a; van Milgen et al., 1998; de Lange et al., 2006). In NRC (2012), MEm is estimated as 197 kcal/ BW0.60 for growing-finishing pigs. However, for gestating and lactating sows, MEm is estimated as 100 and 110 kcal/ BW0.75, respectively.
It is more challenging to estimate NEm than to estimate MEm, but NEm may be calculated as the sum of FHP and energy used for physical activity (van Milgen et al., 2008). However, NEm has also been assumed to be equal to FHP (de Goey and Ewan, 1975; Ewan, 2001; Kil, 2008). Estimates of FHP for growing pigs are between 71 and 81 kcal/ BW0.75 (Robles and Ewan, 1982; Just et al., 1983). If 0.60 is used as the exponent, however, estimates for FHP range from 117 to 179 kcal/BW0.6 (Schiemann et al., 1972; Just et al., 1983; Noblet et al., 1994a; van Milgen et al., 1998; de Lange et al., 2006). The value of 179 kcal/BW0.6 has been widely used for calculating NE values for diets and feed ingredients fed to growing and finishing pigs (Kil et al., 2011, 2013; Stewart et al., 2013; Whittemore et al., 2003). Recently, the following equations to predict daily FHP and NEm for growing and finishing pigs were suggested (van Milgen et al., 2008):
It was assumed that the energy expenditure from daily physical activity of pigs is 207 kJ NE/BW0.6/d. The maintenance adjustment value is set to 1.0 unless pigs are inactive or immune-challenged.
Factors affecting the energy requirement for maintenance
Variations in the maintenance requirement among pigs result from differences in age, sex, genotype, physiological state, environment, and feeding strategy (Baldwin, 1995). Specific conditions such as thermoregulation, immune system activation, disease, and stress also influence the maintenance requirement to a large extent (Bray et al., 1997; Knap, 2000; Verstegen, 2001).
Ambient temperature is a major factor in determining maintenance energy requirements. The FHP of growing pigs is reported to be 16% less at 33°C than at 23°C (van Milgen and Noblet, 2000). Likewise, pigs have 4% greater energy requirement for maintenance for each 1°C reduction of the temperature when they are kept below the lower critical temperature because of increased heat production for maintaining body temperature (Close, 1996). The maintenance requirement may also change if the temperature increases above the evaporative critical temperature as a consequence of increased respiration rate. However, the energy cost for panting is relatively small and a reduction in feed intake decreases the metabolic rate, and hot temperature appears to have little impact on the energy requirement for maintenance (Black, 1995; Giles et al., 1998).
The size of the gastrointestinal tract (GIT), liver, kidney, and heart as well as the relative ratio of body protein to body lipids also impacts the energy requirement for maintenance (Koong et al., 1983; van Milgen et al., 1998; Knap, 2000). Total heat production from metabolically active organs accounts for 25 to 30% of basal heat production (Baldwin and Bywater, 1984). Body organs and the GIT contribute 3 to 4 times more to FHP than skeletal muscles, whereas lipids in the body may decrease FHP or have little impact on FHP (Tess et al., 1984; van Milgen et al., 1998; Noblet et al., 1999). Genotype, sex, feeding strategy, and stage of growth influence the size of metabolically active organs and body composition, and therefore, may result in variation in the energy requirement for maintenance (Tess et al., 1986; Baldwin, 1995; Noblet et al., 1999).
Indirect calorimetry is often used to measure FHP. Estimates of FHP, however, vary with the period of fasting, the type and amounts of previously fed diets, and the physical activity of animals (van Milgen et al., 1998; Ewan, 2001; de Lange et al., 2006). Short-term fasting may better represent the FHP of growing animals because animals may adapt FHP by responding to the feeding level (Koong et al., 1983; van Milgen et al., 1998; Noblet et al., 1999). Restriction of the physical activity of animals in metabolism cages may decrease FHP and group-housed pigs have a greater heat production than individually-housed pigs (van Milgen and Noblet, 2000).
ENERGY VALUES FOR FEED INGREDIENTS AND DIETS
Pigs do not utilize all the energy in the diets, but a direct quantification of energy used for various body functions in pigs is almost impossible. As a consequence, available energy in diets fed to pigs is indirectly determined by subtracting sequential energy loss (i.e., unavailable energy to pigs) in feces, urine, gases, and heat increment during nutrient metabolism from total energy in the diet.
Gross energy
The total energy in feed ingredients is called GE, which represents the maximum quantity of energy (or heat of combustion) that is present in an ingredient and is obtained from the complete combustion of organic materials using bomb calorimetry. The total quantity of GE in an ingredient depends on the degree of oxidation of organic components and is determined by the ratio of C and H to O in the ingredient (McDonald et al., 2002). Dietary nutrients have average GE values of 3.7 kcal/g for glucose, 4.2 kcal/g for starch, 5.6 kcal/g for protein, and 9.4 kcal/g for lipids, whereas fermentative products have GE values of 3.5, 5.0, 5.9, and 13.1 kcal/g for acetate, propionate, butyrate, and methane, respectively (McDonald et al., 2002). Thus, the GE in a feed ingredient can be estimated if the chemical composition of the ingredient is known. However, the GE is totally independent of the animals and provides no indication of available energy to pigs.
Digestible energy
The DE of feed ingredients is calculated from the difference between GE in the diet and GE excreted in the feces. For DE in diets, therefore, energy that is consumed by the animal and is not excreted in the feces is assumed to be absorbed and available to pigs (de Lange and Birkett, 2005). The DE is an apparent measurement of the energy value for the diet because the endogenous losses of energy are disregarded in its calculation (Reynolds, 2000). The DE of most diets fed to pigs varies between 70 and 90% of GE in the diet (Sauvant et al., 2004). In typical corn-soybean meal diets, an average of 82% of the GE in the diet is absorbed and present as DE if fed to growing-finishing pigs (Kil, 2008). The variation in the average utilization of GE results from variations in inclusion levels and sources of fiber, lipids, and minerals in the diets, the level of feed intake, and feed processing techniques (Just, 1982; Noblet, 1996; Black, 2000; Verstegen, 2001). Among these factors, dietary fiber is likely the primary reason for variation in DE values among diets (Noblet and van Milgen, 2004). Animal factors such as age or BW may also influence the DE of diets, probably due to differences among young pigs and older pigs in their capacity to ferment dietary fiber (Shi and Noblet, 1993; Noblet and van Milgen, 2004). In general, DE of diets and feed ingredients increases with BW of pigs as the GIT becomes developed (Noblet and van Milgen, 2004; Kil et al., 2011, 2013).
Metabolizable energy
The ME of a diet is calculated by subtracting energy excreted in urine and gases from DE. However, the gaseous energy losses have often been ignored in the calculation of ME because relatively small amounts of gasses are produced by pigs and because of the difficulty of measuring these losses. Therefore, the difference between DE and ME is usually calculated as the difference between DE and urinary energy excretion. Energy losses in urine depend on urinary N concentration (Noblet and van Milgen, 2004). On average, 50% of absorbed N is used for body protein synthesis and the remaining 50% of absorbed N is excreted in the urine (Just, 1982; Sauvant et al., 2004). However, the ME value only accounts for energy losses as urinary excretion, but do not consider the energy cost for urinary excretion (e.g., the energy cost of synthesizing urea; Birkett and de Lange, 2001).
Recently, increasing use of high fiber ingredients for diets fed to pigs has increased the importance of gaseous energy losses when ME of feeds is calculated because increased dietary fiber results in increased hindgut fermentation and, therefore, increased synthesis and loss of methane. As a consequence, the ME of diets varies with the quantities and characteristics of dietary fiber (Noblet, 1996; NRC, 1998). Energy lost in gasses is reported to range from 0.1 to 3.0% of DE (Shi and Noblet, 1993). Rijnen (2003) proposed the following equation to predict gaseous energy loss as CH4 based on the quantities of fermentable fiber.
Energy that is consumed by animals and is not excreted in the feces, urine, or gases is assumed to be available to the animal (de Lange and Birkett, 2005). Therefore, the ME has been considered a better estimate of available energy to pigs than DE because ME accounts for urinary energy losses and even gaseous energy losses (Just, 1982; Moehn et al., 2005). In growing pigs, the average ratio of DE to ME in commercial diets is approximately 0.96 (Noblet et al., 1994a; NRC, 1998). As observed with DE, however, ME values are also affected by nutrient composition of diets and BW of pigs.
A correction for the ME to N equilibrium (N corrected metabolizable energy, MEn) has been proposed to eliminate the impact of N retention on ME:DE (Morgan et al., 1975). Values for MEn have been suggested to be a better estimate of available energy from protein sources in feeds than ME (Morgan et al., 1975; Robles and Ewan, 1982; de Lange and Birkett, 2005). For calculation of MEn for pigs, 9.17 kcal of ME per g of N retention is subtracted from the measured ME values (Morgan et al., 1975), based on the assumption that for each 1 g of retained N, 9.17 kcal of energy is lost in urine. However, MEn values have not been widely used in pig diets because growing pigs do not usually use the protein retained in the body as an energy source. More recently, however, it was proposed that ME values be standardized to 50% N retention because on average, 50% of the absorbed N is used for protein synthesis (Cozannet et al., 2010).
Net energy
Values for NE are calculated by subtracting the heat increment, which is energy losses during nutrient metabolism, from ME. However, heat increment is difficult to separate from total heat production, and therefore, NE is typically calculated as the sum of NEm and retained energy (Noblet, 2007). The NE value can also be calculated from DE or ME if the efficiency of ME utilization for NE (NE:ME) or that of DE utilization for NE (NE:DE) is known (Baldwin and Bywater, 1984; de Lange, 2008). The ratio of NE to ME represents the energetic efficiency of ME for maintenance and production, and thus depends on the particular biochemical pathways for the purpose of utilization (e.g., maintenance vs. energy retention) in animals (Black, 2000). Theoretical values for NE:ME have been proposed for energetic efficiencies of digestible nutrients for ATP production and lipid retention (Black, 1995; Table 1). The NE:ME of digestible nutrients is 58, 82, and 90% for protein, starch, and lipids, respectively, and the average for the NE:DE and NE:ME of feeds is 71 and 74%, respectively (Noblet et al., 1994a). Because NE values are obtained based on the energetic efficiency of digestible nutrients, NE values for diets and feed ingredients more closely represent the available energy than DE or ME values (Noblet et al., 1994a; Moehn et al., 2005; Noblet, 2007). An example of approximate utilization of energy in diets is presented for growing pigs fed corn-soybean meal-based diets (Kil, 2008; Figure 2).
Although NE values have been recognized as better estimates for available energy in diets and feed ingredients for pigs than DE and ME values, there is a concern that more accurate descriptions of available energy in diets increases the dependency on individual pigs, which increases variations in energy values for diets because of variations among pigs (Boisen and Verstegen, 2000). One of the concerns is the determination of NEm, which is used for the NE calculation and varies greatly among pigs due to variations among individual animals and the environment where the animals are located (Baldwin and Bywater, 1984). In theory, an NEm value measured under specific conditions of an animal cannot be applied to an animal kept under different conditions and for animals with different characteristics (Boisen, 2007). This problem was illustrated when NEm was estimated for both growing and finishing pigs kept at 3 different locations in North America. Results of this experiment indicated that values for NEm, which were estimated using the regression method (i.e., extrapolated FHP) were different among locations demonstrating significant effects of environmental conditions on measured values for NEm. It was also demonstrated that the growth stage of pigs influenced determined values for NEm (Kil, 2008; Table 2). However, most previous experiments have used 1 constant value for NEm (e.g., 179 kcal/kg BW0.6) in the calculation of NE values, and applied this value to all categories of pigs. This approach may be acceptable in calculating NE values of feed ingredients for the use in feed evaluation systems because the impact of using different values for NEm will only change the actual values for NE, but will not influence the ranking among feed ingredients. However, if the actual NE of a diet fed to a group of pigs raised under a specific environment is to be determined, it is likely that an inaccurate estimate for NE may be obtained if a constant pre-determined value for NEm is applied for the NE calculation.
Another concern with the NE system is that the efficiency of ME for NE (ME:NE) changes with stage of growth of pigs. For instance, finishing pigs deposit more lipids than growing pigs (de Greef et al., 1994; Kil et al., 2011, 2013; Stewart et al., 2013) and the energetic efficiency of ME for lipid retention is greater than for protein retention or body maintenance (Black, 1995). Therefore, the NE of a diet appears to be greater for finishing pigs than for growing pigs if the same diet is fed to both growing and finishing pigs. This observation is supported by recent experiments reporting the NE of feeds and various feed ingredients (e.g., corn, soybean oil, soybean hulls, and wheat middlings) in growing and finishing pigs (Kil et al., 2011, 2013; Stewart et al., 2013). The implication of these observations is that NE values for diets and feed ingredients are not constant across different stages of growth for pigs. In an attempt to avoid this problem, NRC (2012) proposed a new concept of effective ME. The effective ME of diets is calculated from the NE of feeds with specific conversion factors that represent the NE:ME of feeds for each stage of growth. The different conversion factors are assigned to starting pigs (5 to 25 kg; 1/0.72) and growing-finishing pigs (25 to 135 kg; 1/0.75). However, this approach does not account for differences in NE:ME of diets between growing and finishing pigs, and the applicability of using this approach for pig diets has not been tested yet. It may also be argued that the NE of diets changes during the growing-finishing period from 25 to 135 kg as has recently been demonstrated (Kil et al., 2011, 2013; Stewart et al., 2013), and therefore, those different NE:ME ratios should be applied during this period to further improve the accuracy of the calculated values for effective ME.
The methodology used to measure energy retention also influences calculated NE values. The comparative slaughter method and the indirect calorimetry method have been used to measure energy retention or total heat production for pigs. Each method has advantages and disadvantages. The comparative slaughter method provides a better estimate of body composition gain and may more closely simulate practical feeding systems than the indirect calorimetry method. However, measurements of body energy retention may be associated with inherent errors because the initial energy concentrations of experimental pigs are predicted from a different group of pigs that are slaughtered at the initiation of the experiment (Quiniou et al., 1995). To minimize variation in body chemical composition among animals, relatively long experimental periods and a large number of animals in the initial slaughter group, which have similar genetic background as pigs fed the experimental diets, are required (Boisen and Verstegen, 2000).
The indirect calorimetry method has the advantage of measuring energy retention with the same pigs during a short period of time. However, pigs are often placed in confined respiratory chambers, which may reduce the physical activity and thermoregulation response (Baldwin and Bywater, 1984; Reynolds, 2000). Direct extrapolation of values obtained under these conditions to practical feeding systems may not always be appropriate. It has been reported that estimates of energy retention tend to be greater if measured by the indirect calorimetry method than by the comparative slaughter method, which directly influences the calculated values for NE of the diet (Quiniou et al., 1995; van Milgen and Noblet, 2003; Kil et al., 2011, 2013).
ENERGY SYSTEMS
An energy system is defined as a way of assigning energy values to feed ingredients using prediction equations. These predicted energy values are supposed to be additive in mixed diets and are used to calculate the energy concentrations in mixed diets (Emmans, 1999). An appropriate energy system is also able to accurately estimate the amounts of energy required for maintenance and production, and to correctly rank energy values in a variety of feed ingredients (Noblet et al., 1994a; Rijnen et al., 2004). Therefore, the quality of an energy system is dependent on the accuracy with which the system can predict animal performance (Noblet and Henry, 1993).
DE and ME systems
The DE and ME systems have been the most widely used for evaluating ingredients and diets fed to pigs because values for DE and ME are relatively easy to measure and these values are assumed to be additive in mixed diets (NRC, 1998; de Lange and Birkett, 2005). The DE and ME systems are compatible because DE values can be readily converted to ME values, assuming that the average efficiency (ME:DE) is 0.96 (Noblet et al., 1994a; NRC, 1998). However, the ME:DE varies with BW (Noblet et al., 1994b), the amount and quality of protein (Whittemore, 1997), and the amount and characteristics of fermentable fiber in the diets (Le Goff and Noblet, 2001). Values for DE and ME are additive in mixed diets only if they are measured in pigs at the same stage of growth and fed a similar amount of diet (Rijnen et al., 2004). It is suggested that the DE and ME systems may underestimate energy values for ingredients high in lipids and starch, but overestimate energy values for ingredients high in protein and fiber (Noblet et al., 1994a; Noblet, 2007), because ingestion of these nutrients results in different quantities of heat increment (Noblet et al., 1994a; Black, 1995). Therefore, DE and ME systems may have limitations in accurately predicting the energy cost for growth of pigs (Le Bellego et al., 2001; Rijnen et al., 2004; Wu et al., 2007).
Several equations predicting the DE and ME of ingredients and diets for growing pigs have been proposed:
DE ( kcal / kg ) = 4 , 168 − ( 91 × % Ash ) + ( 19 × % CP ) + ( 39 × % EE ) − ( 36 × % NDF ) ; NRC ( 2012 ) adapted from Noblet and Perez ( 1993 )
Energy and chemical components are expressed on a DM basis in all equations.
NE system
The NE system has been used in several European countries to overcome the limitations of DE and ME systems (de Lange and Birkett, 2005). The assumed advantages of NE over DE and ME include a better prediction of growth performance and body composition of animals (Verstegen, 2001; Noblet, 2007; Oresanya et al., 2008). Wu et al. (2007) reported that the energy cost per unit of BW gain (kcal/kg BW gain) was constant for growing-finishing pigs fed diets containing increasing amounts of fat when dietary NE is used for the calculation, whereas it linearly decreased when dietary DE or ME is used for the calculation, which indicates that the NE system is superior to DE or ME system in terms of predicting actual energy cost required for BW gain. This advantage is more obvious if a diet contains several different feed ingredients than if the diet contains primarily corn and soybean meal (Le Bellego et al., 2001; Patience and Beaulieu, 2005; de Lange, 2008). The utilization of N may also be improved because diets can be more accurately formulated with the proper ratio of amino acids to energy (i.e., NE), which leads to a reduction in waste excretion (Le Bellego et al., 2001; Payne, 2006; Noblet, 2007). The NE system may also reduce diet costs if this system can be used to more correctly rank feed ingredients in terms of energy values (Patience and Beaulieu, 2005; Payne, 2006).
The NE systems in France and the Netherlands were developed using similar principles to predict NE values for mixed diets. Using these systems, it is assumed that the chemical composition and nutrient digestibility of feed ingredients can be obtained from tabulated values, that the amounts of digestible nutrients in each ingredient are additive in mixed diets with no interactions among feed ingredients, and that the utilization of a digestible nutrient for energy in the body is independent of the other digestible nutrients (Bakker, 1996). Prediction equations for the NE of feed ingredients are established using data obtained from the chemical composition and digestibility of nutrients.
The French NE system
The French NE system was developed by Noblet and his coworkers who conducted numerous experiments at the Institut National de la Recherche Agronomique (INRA). In short, NE values of 61 diets containing a variety of feed ingredients at a high ME intake (550 kcal of ME/kg BW0.6) or at a low ME intake (330 kcal of ME/BW0.6) were determined with lean-genotype growing pigs (35 kg of initial BW; Noblet et al., 1994a). Energy digestibility and energy losses in feces, urine, and gases were measured in respiratory chambers. Heat production was measured using the indirect calorimetry method. The average daily FHP for growing pigs was determined as 179 kcal/kg BW0.6 by extrapolating total heat production to zero ME intake using regression analysis. The NE values of each diet were then calculated as the sum of FHP and energy retention in pigs fed at high ME intake. Eleven regression equations based on the concentrations of digestible nutrients and dietary nutrients were proposed for predicting NE values in mixed diets and feed ingredients (Noblet et al., 1994a). Components of digestible nutrients in the equations were later modified and 3 equations were then used (Sauvant et al., 2004). It is assumed that these 3 equations are applicable to both mixed diets and feed ingredients for growing-finishing pigs (Noblet and van Milgen, 2004; Sauvant et al., 2004). The 3 equations are the following:
In all equations, energy and chemical components were expressed on a DM basis; digestible residue = digestible OM-digestible CP-digestible EE-starch-sugar. More recently, NRC (2012) suggested 3 equations that are slightly modified from the French NE systems.
The Dutch NE system
The Central Bureau Livestock Feeding (CVB) has developed the Dutch NE system. This system uses the concentrations of digestible nutrients in feed ingredients to estimate the NE values of feeds and feed ingredients in a way that is consistent with the French NE system (Rijnen et al., 2004). However, the Dutch NE system separates total digestible carbohydrates (i.e., starch and sugar) into an enzymatically-digestible fraction and a fermentable fraction because of differences in energetic utilization of carbohydrates between the small intestine and large intestine of pigs (Blok, 2006). Starch is analyzed using an enzymatic procedure with amyloglucosidase rather than with the Ewers polarimetric procedure as is used in the French system – this difference results in differences in analyzed values for starch in the ingredients. Sugars are analyzed for the enzymatically degraded fraction of the total sugars and the remaining is considered fermentable sugars. Digestible lipids are analyzed using the acid hydrolyzed ether extract (AEE) method for both feeds and feces, whereas in the French NE system, lipids are determined using acid hydrolyzed ether extract method only for feces. The NE of diets and feed ingredients are calculated in the Dutch system using the following equation (Blok, 2006):
where energy and chemical components are expressed on a DM basis; starch-e = enzymatically digestible starch, sugar-e = enzymatically digestible sugar, FCH (fermentable carbohydrates) = fermentable starch (starch-f, zero value except for potato starch)+fermentable sugar (= total sugar – sugar-e)+digestible NSP; digestible NSP = digestible OM − digestible CP − digestible EE − starch-e − 0.95×total sugar.
The Danish potential physiological energy (PPE) system
The Danish energy evaluation system is unique because a new concept called “Potential Physiological Energy (PPE)” is used. The system is based on the theoretical biochemical utilization of energy (i.e., ATP) by pigs. Therefore, it is closer to a mechanistic model than to other NE systems. The value for PPE of nutrients is the potential energy value for ATP production if digestible nutrients are completely oxidized by animals (Boisen, 2007; Table 3). The PPE of different nutrients is assumed to be independent of their metabolic utilization (e.g., oxidation or retention), and therefore, the PPE calculated from various feed ingredients or digestible nutrients are additive in diets containing a mixture of feed ingredients and are independent of animal factors (Boisen, 2007). All data for concentrations of digestible nutrients in feed ingredients can be obtained from tables for feed ingredients. The Danish PPE system also uses in vitro digestibility procedures to estimate the digestibility of CP, amino acids, OM, lipids, and carbohydrates to avoid the effects of animals on nutrient digestibility. An estimate of enzyme-undigested DM at the distal ileum is also obtained from the in vitro procedure to correct the energy value for the compounds originating from endogenous synthesis of protein and lipids throughout the GIT. The energy values for feeds in this system are expressed as Feed units (FU), which are calculated from the PPE values of each nutrient in the diet. One feed unit for growing pigs is equivalent to 7.38 MJ PPE. The following equation is used to estimate the energy in this system:
where FU is expressed on a DM basis and other components are based on g/kg DM; RDCP = in vitro ileal digestible CP, RDCF = calculated ileal digestible fat, EDC = in vitro ileal digestible carbohydrates, FERMC = fermentable carbohydrates, EIDMi = enzyme−undigested ileal DM.
Comparison of European energy systems
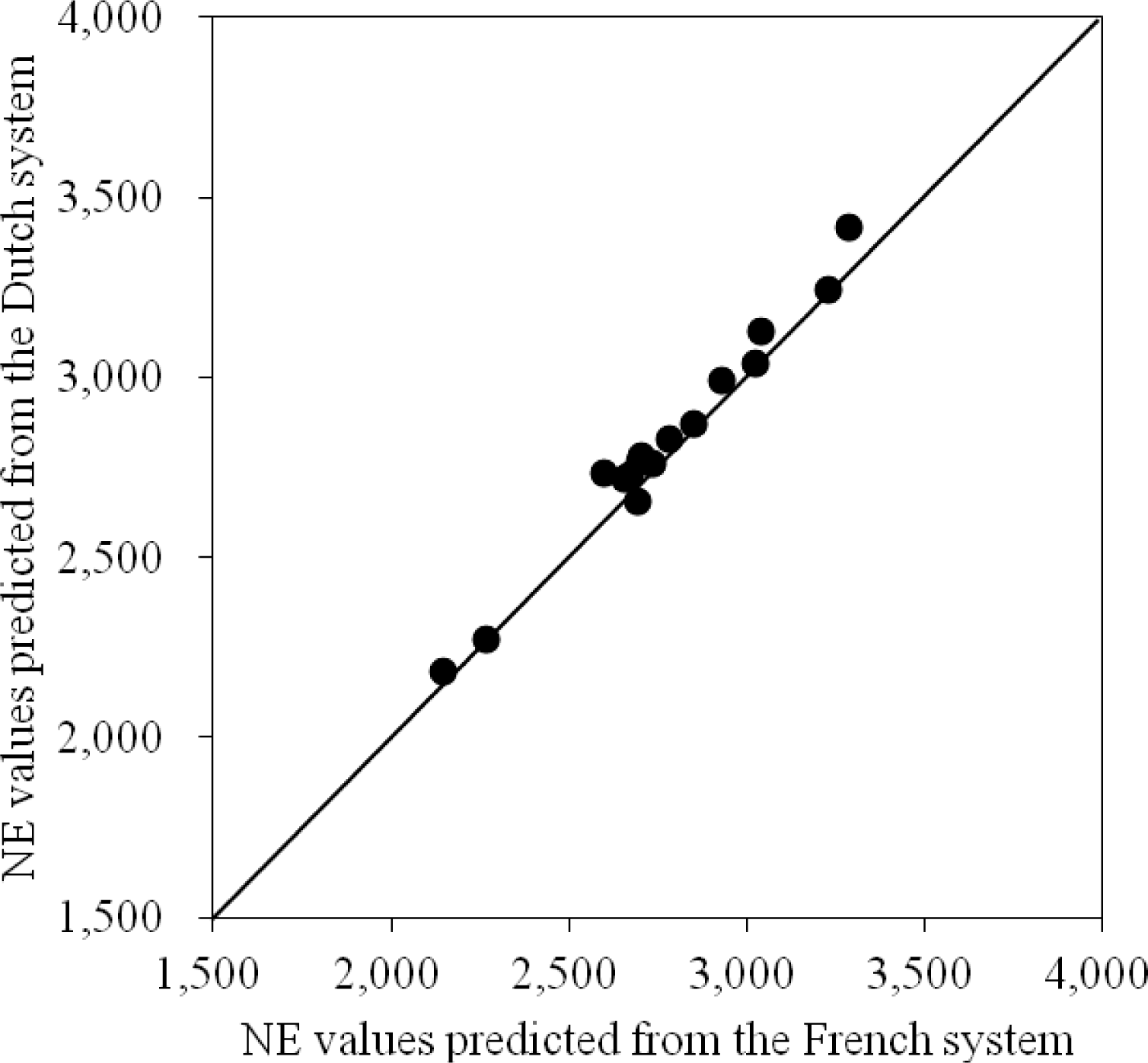
The NE values for diets and feed ingredients are affected by energy evaluation systems (Noblet and van Milgen, 2004). The accuracy and applicability of an energy system are largely influenced by the relative contributing energy values (i.e., assigning factors) for different digestible nutrients (Boisen, 2007; Table 4). Starch is used as the standard reference nutrient for estimating energy contributions among nutrients because starch is considered a major energy source for pigs (Boisen, 2007). The energy contribution of nutrients relative to starch among European energy systems is presented in Table 5. The relative contribution of different digestible nutrient fractions seems comparable among these energy systems. Therefore, the hierarchy of energy values for diets and feed ingredients is expected to be similar among the 3 European energy systems although absolute energy values may differ. Kil (2008) compared the predicted NE values from the French and the Dutch system using 16 mixed diets containing various feed ingredients. In general, the values predicted from the Dutch system appeared to be slightly greater than those predicted from the French system; however, the hierarchy of NE values of the 16 diets as affected by different nutrient compositions was close between these 2 NE systems (Figure 3).
CONCLUSIONS
Energy in diets is digested, absorbed, and metabolized by pigs and subsequently used for maintenance or productive purposes. Accurate estimation of energy values in feed ingredients makes it possible to formulate diets that ensures optimal growth of pigs and minimizes feed costs. Different energy systems (DE, ME, NE, or PPE system) have been developed and used to quantify the amounts of available energy in feed ingredients and diets. The NE system appears to be superior to DE or ME systems if diets contain ingredients high in protein or fiber because the NE system corrects for energy lost as heat increment during digestion and nutrient metabolism. The PPE system may also be superior to DE or ME systems because this system is considering only the energy that can be potentially used by the animal. However, none of the energy systems are able to accurately predict true energy values of diets due to significant interactions among animals, the environment, and feed ingredient characteristics. In the future, therefore, it is possible that improved energy systems will be developed. Such systems will likely be based on sophisticated analytical procedures for nutrients in feed ingredients and mechanistic modeling that describes energy transformations for body functions as affected by animals, environment, and feed ingredient characteristics.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







