



INTRODUCTION
Hen egg yolk phosvitin is a phosphoglycoprotein that forms the structure of yolk granules by making phosphocalcic bridges with high-density lipoprotein (HDL) (Taborsky, 1983). It represents about 7% of the yolk protein (Mecham and Olcott, 1949). Phosvitin is composed of α- and β-phosvitin. The α-phosvitin is an aggregate of three or four sub-units of 35 to 40 kDa and the β-phosvitin is an aggregate of four or five subunits of 45 kDa (Anton et al., 2007). These have a characteristic amino acid composition, which is approximately 50% serine, and most of which are phosphorylated (Clark, 1985). This specific structure makes phosvitin a potent metal chelator. Typically, phosvitin isolated from chicken egg yolk contains 2 to 3 atoms of iron per molecule and two moles of organic phosphorus from phosvitin can bind one mole of iron (Mecham and Olcott, 1949; Taborsky, 1963). Thus, a phosvitin molecule can bind up to 65 to 70 iron molecules under optimal conditions (Taborsky, 1963; Webb et al., 1973).
The metal chelating ability of phosvitin enables it to be used as a functional material. Ishikawa et al. (2004) found that phosvitin and its proteolytic hydrolysates obstruct the generation of iron-catalyzed hydroxyl radicals from the Fenton reaction. And, many researchers have reported that phosvitin inhibited iron-catalyzed lipid oxidation in meat (Lee et al., 2002; Jung et al., 2012a; Jung et al., 2013), and exhibited an antibacterial effect on iron deficient bacteria (Khan et al., 2000; Choi et al., 2004). Additionally, Jung et al. (2012b) suggested phosvitin as melanogenesis inhibitor. For these reasons, phosvitin has received attention, and many studies have attempted to separate phosvitin from chicken egg yolk.
Phosvitin extraction and purification methods reported in previous studies used organic solvents, such as ether, a combination of chloroform and methanol, a combination of hexane and ethanol, or ethanol to remove lipids and phospholipids from yolk granules (Heald and McLachlan, 1963; Wallace and Morgan, 1986; Losso and Nakai, 1994). Although these methods separate phosvitin from chicken egg yolk, the extracted phosvitins have limited use because of the solvents used, and these methods are relatively complicated. Furthermore, some of the solvents used can modify phosvitin structure or decrease the recovery of phosvitin (Castellani et al., 2003; Ko et al., 2011). Improved methods for separating phosvitin from chicken egg yolk have been introduced recently. Highly purified phosvitin (98%) with a high recovery rate (85%) was produced by an isolation method based on the insolubility of Mg2+/phosvitin salt followed by ion-exchange chromatography (Castellani et al., 2003). Also, phosvitin was successfully separated using ethanol and NaCl with a 96% recovery rate (Ko et al., 2011). These two methods can be used to develop large-scale production of phosvitin, but these methods still use high priced equipment or organic solvents.
Thermal precipitation is a precipitation technique used to isolate and purify proteins. Proteins denatured by thermal treatments can be removed by mild centrifugation (Burgess, 2009). Yolk granules contain 70% high density lipoprotein (HDL), 16% phosvitin, and 12% low-density lipoproteins (LDL) with different thermal stabilities (Anton et al., 2000). Phosvitin and β-HDL are not affected by thermal treatments up to 82°C, whereas LDL and α-HDL are denatured at 72 and 76°C, respectively. Phosvitin can be extracted from yolk granules by disintegrating granule structure using 0.5 M NaCl (Le Denmat et al., 1999; Anton et al., 2000). Additionally, the extracted phosvitin does not aggregate and iron binding ability is not influenced when the solution is heated at 90°C for 60 min (Castellani et al., 2004).
Therefore, we hypothesized that phosvitin could be extracted with salt and purified by thermal treatment without using organic solvents. The phosvitin separation procedure using thermal treatment is simpler than previously developed methods and appropriate for large-scale phosvitin production. The objective of this study was to develop a new separation method for phosvitin from egg yolk without using organic solvents.
MATERIALS AND METHODS
Chemicals
Chicken eggs were obtained from a local market (Daejeon, Korea). Phosvitin standard, acrylamide, N,N’-methylene-bis-acrylamide, tris (hydroxymethyl) aminomethane (Tris), sodium dodecyl sulfate (SDS), ammonium persulfate, N,N,N’,N’-tetramethyle-ethylenediamine (TEMED), glycine, glycerin, Coomassie Brilliant Blue G-250, aluminum nitrate, and Tween 20 were obtained from Sigma-Aldrich (St. Louis. MO, USA). Protein 2× sample buffer was obtained from Elpis-Biotech (Daejeon, Korea). Sodium chloride, potassium chloride, and ammonium sulfate were obtained from Junsei Chemical (Tokyo, Japan). Enhanced chemiluminescence (ECL) plus Western blotting detection reagent was obtained from GE Healthcare (Buckinghamshire, UK). Mouse monoclonal IgG2a and goat anti-mouse IgG-horseradish peroxidase were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Prestained protein markers were obtained from iNtRon Biotechnology (Sungnam, Korea).
Phosvitin extraction
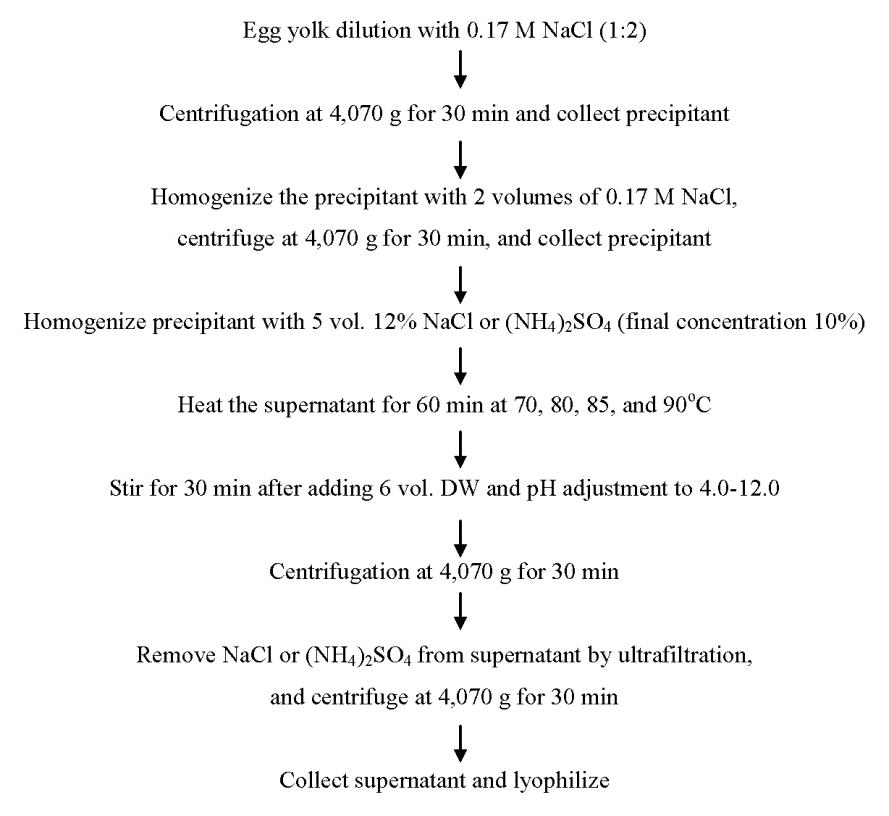
Yolk was separated from egg white, and carefully rolled on a paper towel to remove all egg white. Then, the egg yolk membranes and chalaza were removed by filtering through a 2,000 μm testing sieve (Chung Gye Sang Gong Sa, Seoul, Korea). The filtered egg yolk was homogenized with two volumes of cold 0.17 M NaCl, and centrifuged at 4,070×g for 30 min at 4°C. The precipitant was collected and re-suspended in two volumes of cold 0.17 M NaCl, homogenized, and centrifuged at 4,070×g for 30 min. The precipitant containing yolk granules was collected, and homogenized with five volumes of 12% NaCl or 12% (NH4)2SO4 (final concentration was 10%). The disrupted granules solution was subjected to thermal treatment in a water bath at 70, 80, 85, or 90°C for 1 h. After heating, the sample solutions were diluted (1:6, v/v) with distilled water, and the pH of the sample solutions was adjusted to pH 4.0 to 12.0 using 6 N HCl or 10 N NaOH. After stirring for 30 min, the homogenates were centrifuged at 4,070×g for 30 min at 4°C. The supernatant was filtered through filter paper (Whatman No.1) to remove floating particles. The filtrate was concentrated and desalted using an ultrafiltration system (Quixstand Benchtop System using a membrane column with a 10 kDa molecular weight cut-off; GE Healthcare, Waukesha, WI, USA), and then centrifuged at 4,070×g for 30 min at 4°C. The supernatant was collected and lyophilized (Figure 1).
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE)
SDS-PAGE was performed using a 10% polyacrylamide gel containing 30% acrylamide solution, 1.5 M Tris-HCl (pH 8.8), 0.5 M Tris-HCl (pH 6.8), 10% ammonium persulfate, and TEMED. The extracted protein solution was mixed with the same volume of 2× sample buffer composed of 125 mM Tris-HCl (pH 6.8), 20% glycerol, 2% SDS, 2% mercaptoethanol, and 0.02% bromophenol blue, and heated at 95°C on a heating block for 2 min. Pre-stained protein markers (9 to 198 kDa) were used as molecular markers. Electrophoretic separation was performed with the pageRun system (AE-6531 mPAGE, ATTO Co., Tokyo, Japan) by applying 40 mA for 40 min. The running buffer was composed of 25 mM Tris, 0.1% SDS, and 192 mM glycine. Proteins in the gels were stained with Coomassie Brilliant Blue containing 0.1 M aluminum nitrate, and then destained in a 10% acetic acid solution. The stained gel was scanned using a GS-710 (Bio-Rad Laboratories Inc, Hercules, CA, USA) densitometer at an optical resolution of 63.5 μm/pixel and then analyzed by Image Master 2D Platinum v5.0 (GE Healthcare, formerly Amersham Biosciences). The relative phosvitin composition in the protein solution was estimated from band intensity (pixel intensity×band area).
Iron-binding ability of phosvitin
The iron-binding ability of the isolated phosvitin was determined using a modified ferrozine method (Riemer and Hoepken, 2004). A mixture of 3.3 mL distilled water, 50 μL 3.0 mM FeCl3 or 3.0 mM FeCl2.4H2O in distilled water, and 50 μL phosvitin solution were placed in a 15 mL conical tube. The mixture was left at room temperature for 1 min. Next, 0.4 mL of 5 mM ferrozine was blended with 0.2 mL 1% ascorbic acid in distilled water and added to the tubes. After 5 min incubation at room temperature, the final color change was monitored using a spectrophotometer (DU530, Beckman Instruments Inc., Fullerton, CA, USA) at 562 nm using distilled water as a blank. Iron binding ability was calculated with the following equation:
Where, A0 indicates the absorbance of the sample solution without phosvitin and A1 indicates the absorbance of the sample solution with phosvitin.
Western blotting
Proteins in the SDS-PAGE gel were transferred to pure nitrocellulose membranes (Trans-Blot Transfer medium; Bio-Rad) using the Trans-Blot apparatus (Bio-Rad). The transfer buffer was composed of 24 mM Tris, 192 mM glycine, and 20% (v/v) methanol. After transfer, the membranes was blocked with 5% nonfat dry milk (w/v) in TBS-T solution (pH 7.4) composed of 24.8 mM Tris, 137 mM sodium chloride, 2.7 mM potassium chloride, and 0.05% Tween 20 for 1 h at room temperature. After blocking, the membrane was washed six times (10 min/wash) with TBS-T and then incubated at room temperature for 1 h with phosvitin primary antibody (mouse monoclonal IgG2a, Santa Cruz Biotechnology) diluted with TBS-T at the ratio of 1:2,500. The membrane was rewashed three times (10 min/wash) with TBS-T for removing primary antibody and then incubated at room temperature for 1 h with the secondary antibody (goat anti-mouse IgG-horseradish peroxidase, Santa Cruz Biotechnology) diluted with TBS-T at the ratio of 1:10,000. After the secondary antibody incubation, the membrane was washed three times (10 min/wash), and the detection procedure was carried out in a darkroom using an ECL kit (ECL Plus Western Blotting Detection Reagents, RPN2132, GE Healthcare). The detection reagent was poured onto the membrane and incubated for 1 min. The detection reagent was drained off, and the membrane was exposed to a sheet of diagnostic film (AGFA, Mortsel, Belgium) in a film cassette for 1 min.
Purity of extracted phosvitin
Phosvitin contains 8 to 10% phosphorus and 6.5% carbohydrates. Thus, phosvitin contains 13% nitrogen (Shainkin and Perlmann, 1971; Guérin-Dubiard et al., 2002). The purity of extracted phosvitin was estimated by multiplying nitrogen content X 7.69. Nitrogen content of the phosvitin was determined by Kjeldahl method with automatic Kjeldahl analyzers (Gerhardt GmbH & Co. KG, Germany).
Statistical analysis
All experimental treatments were performed in triplicate. An analysis of variance was performed on raw data, and the mean values and standard deviations were calculated using Statistical Analysis System software (SAS Institute, Cary, NC, USA). Differences among the means were determined by Duncan’s multiple range test and p<0.05 was considered significant.
RESULTS AND DISCUSSIONS
Separation of egg yolk plasma and granules
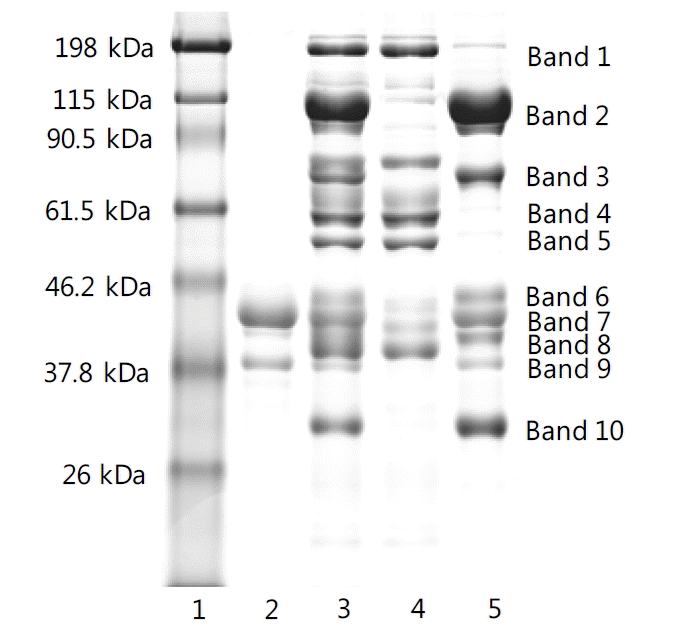
To separate egg yolk plasma and granules, egg yolk was diluted with 0.17 M NaCl and then centrifuged at 4,070 g for 30 min. The supernatant plasma and precipitated granules were diluted in 1% NaCl and delipidated using acetone to clearly identify the protein composition on the SDS-PAGE gel. The SDS-PAGE results are shown in Figure 2. The granules contained LDLs [bands 1 (203 kDa), 4 (62 kDa), and 5 (55kDa)], HDLs [bands 2 (110 kDa), 3 (78 kDa), and 10 (31 kDa)], and phosvitin [bands 7, 8, and 9 (40 to 45 kDa)] which were identical to standard phosvitin (Guilmineau et al., 2005). These results confirmed that phosvitin has several components (α1, α2, and β-phosvitin) and that the major component is β-phosvitin (45 kDa) (Castellani et al., 2003; Ko et al., 2011). However, other studies have reported that the molecular weight of phosvitin is 35 or 55 kDa as a single band (Jiang and Mine, 2000; Guilmineau et al., 2005). A protein (band 6) located near β-phosvitin was not reported in previous studies (Jiang and Mine, 2000; Castellaniet al., 2003; Guilmineau et al., 2005; Ko et al., 2011), but it was thought to be phosvitin because it appeared on the SDS-PAGE results of standard and extracted phosvitin (Figure 7). Several protein bands were identical between plasma and granules. Guilmineau et al. (2005) reported that the protein bands at 203 and 55 kDa in plasma are associated with γ-livetin and a part of α-livetin, respectively. The plasma protein bands at 40 to 45 kDa could be β-livetin (Hatta et al., 2008).
Optimal temperature for extracting phosvitin in the disrupted granule solution
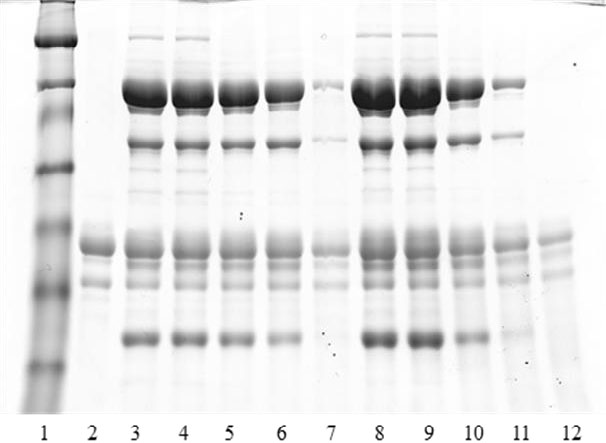
The extracted phosvitin solutions were diluted to decrease the concentration of NaCl or (NH4)2SO4 to approximately 1.5%, because HDLs become insoluble when the ionic strength is <0.3 M NaCl (about 1.8%) (Anton et al., 2007). However NaCl or (NH4)2SO4 was used in the present study at the same concentrations although the ionic strength of (NH4)2SO4 is higher than that of NaCl at the same concentration. The proteins in the supernatants after centrifugation of the diluted solution were separated by SDS-PAGE (Figure 3).
The LDL apo-protein bands disappeared, and the intensity of the HDL apo-protein bands in the extracted solutions decreased gradually as the heating temperature increased to 80°C or higher regardless NaCl or (NH4) 2SO4 treatment. Thermal treatment at 70°C, however, had no effect on denaturing the granular proteins. This result agreed with previous studies suggesting that granular proteins, except for phosvitin and β-HDL, in the disrupted granule solution containing 0.5 M NaCl, were denatured by thermal treatments over 72 and 76°C (Le Denmat et al., 1999; Anton et al., 2000). However, in this study, the intensity of β-HDL bands decreased following thermal treatment at 80°C with (NH4)2SO4, and these bands shrunk remarkably when disrupted granule solutions were subjected to thermal treatment at 90°C with NaCl and 85°C with (NH4)2SO4. Particularly, 90°C thermal treatment of samples extracted with (NH4)2SO4 precipitated all β-HDL from the samples.
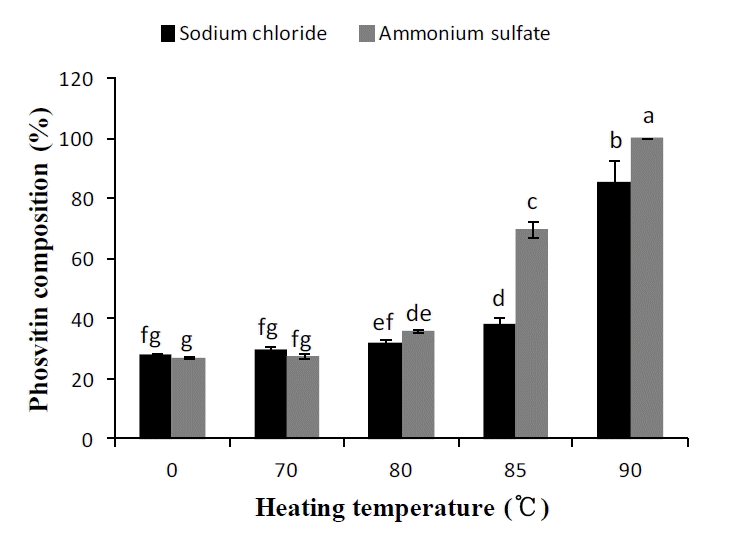
The phosvitin composition among proteins in the extracted solution was expressed as the relative intensity of phosvitin bands in relation to the sum of all band intensity in each lane (Figure 4). The 85°C thermal treatment with (NH4)2SO4 or the 90°C treatment with both NaCl and (NH4)2SO4 significantly increased the proportion of phosvitin among proteins in the extracted solution due to denaturation and precipitation of all other granular proteins (p<0.05). Phosvitin composition was the highest in samples extracted with (NH4)2SO4 and then treated at 90°C when compared with lower thermal treatments or NaCl extracts (p<0.05). The NaCl or (NH4)2SO4 concentrations necessary to disrupt phosphocalcic bridges between phosvitin and HDL were adapted from a previous study (Ko et al., 2011). Ko et al. (2011) suggested that the phosvitin extraction yield from delipidated granules homogenates with 10% (NH4)2SO4 is significantly higher than that with 4, 15, 20, or 25% (NH4)2SO4. Additionally, the purity of the extracted phosvitin using 10% (NH4)2SO4 is significantly higher than that of the 10% NaCl, even if the phosvitin yield is higher when 10% NaCl was used. The present work agreed with the results of Ko et al. (2011), and the difference in purity might be due to the difference in salting-out ability between NaCl and (NH4)2SO4. Ammonium sulfate is a strong salting-out agent for proteins and its salting out ability is higher than that of NaCl (Dumetz et al., 2008).
From the results, the optimal temperature for removing other proteins from the phosvitin extracts was 90°C, and the use of the (NH4)2SO4 for disrupting granules was more effective to remove other proteins from the phosvitin extracts than was NaCl.
Effect of pH on phosvitin solubility in the (NH4)2SO4− extracted solution after the 90°C thermal treatment
Protein solubility is very important for extracting proteins because an increase in protein solubility improves protein yields. Protein solubility in an extracted solution can be increased by adding promoters such as ionic compounds, salts, detergents, and osmolytes (Golovanov et al., 2004).
Phosvitin solubility was expressed as phosvitin composition of the extracted solution (Figure 5). The initial pH of the extracted solution, which contained approximately 1.5% (NH4)2SO4, was about 6.6±0.1. Phosvitin solubility was affected by the pH of the extracting solution. Increasing the pH of the extracting solution above the initial pH significantly increased phosvitin solubility (p<0.05). In contrast, decreasing the pH of the extracting solution below the initial pH significantly decreased phosvitin solubility (p<0.05). Phosvitin solubility was the highest at pH ≥9 and the lowest at pH 4, which is the isoelectric point of phosvitin. However, the β-HDL band started to appear at pH ≥10 on the SDS-PAGE gel (data not shown). Previous studies reported that most egg yolk proteins are soluble in saline buffer (0.56 M NaCl at pH 9), and that phosvitin solubility gradually increases when the pH is increased from 4 to 10 (Guilmineau et al., 2005; Chay Pak Ting et al., 2010). However, Castellani et al. (2003) reported that the highest phosvitin solubility (97.7%) was observed at pH 7 with low MgSO4 concentrations. The reason for the different pH effects on phosvitin solubility could be due to differences in salt concentrations or the kind of salts used.
This result suggested that adjusting pH of the extracting solution to 9 could improve the phosvitin yield, although it required a larger amount of NaOH because of the buffering ability of (NH4)2SO4 at pH >8.
Western blotting
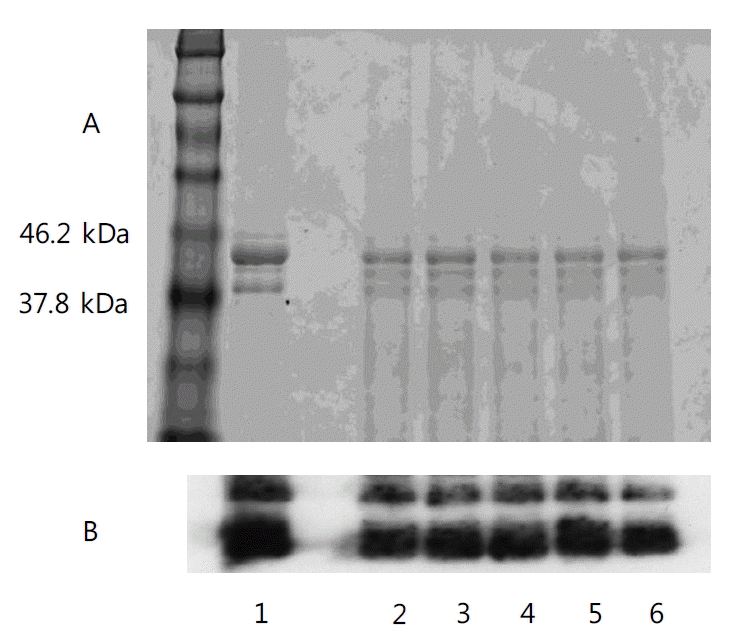
After centrifugation (at 4,070 g for 30 min) of pH 9 phosvitin solution extracted by (NH4)2SO4 with the 90°C thermal treatment, the supernatant was concentrated and desalted using ultrafiltration, and then lyophilized to obtain the final product. The SDS-PAGE result showed that the final phosvitin sample with five repetitions were similar to standard phosvitin in quality (Figure 6). However, a previous study indicated that it is difficult to conclude whether the band is phosvitin using the SDS-PAGE pattern of extracted phosvitin alone, because some proteins in egg yolk have similar molecular weights to phosvitin (Ko et al., 2011). For this reason, the extracted phosvitins were identified by Western blotting. The result of Western blotting confirmed that the extracted phosvitins were identical to standard phosvitin.
Yield, purity, and iron-binding ability of the separated phosvitin
The yields and purities of the extracted phosvitin with five repetitions using this method are shown in Table 1. The average yield of extracted phosvitin from the current method was 56%, and the average purity was 78%. The recovery rate of phosvitin from egg yolk was calculated using the value (12 mg phosvitin/g yolk) reported by Juneja and Kim (1997). In the present study, the granules disrupted by 10% (NH4)2SO4 were heated at 90°C to precipitate other proteins from the granules. As a result, only phosvitin bands are shown in the extracted solution after the 90°C heat treatment (Figure 3). However, the intensity of the phosvitin bands in the extracted solution after heating at 90°C decreased when compared with that after heating at 70°C (data not shown). This may be the consequence of co-precipitation of some of the phosvitin with other proteins, which lead to a lower phosvitin recovery rate.
The extracted phosvitin showed approximately 22.2 and 50.7% ferrous and ferric iron binding ability, respectively, even though these percentages were significantly lower than those of the standard phosvitin (38.2% of ferrous iron and 63.4% of ferric iron) (p<0.05). This result agreed with that of a previous study, which suggested that phosvitin has a higher affinity for ferric iron than ferrous iron (Ishikawa et al., 2004). The iron-binding capacity of phosvitin is about 65 to 70 moles of iron per mole of phosvitin, which can inhibit lipid oxidation catalyzed by the Fenton reaction (Fe2+/H2O2) (Taborsky, 1983; Ishikawa et al., 2004). Ishikawa et al. (2004) suggested that phosvitin has higher antioxidant activity than that of other iron-binding proteins such as transferrin and ferritin. Therefore, extracted phosvitin could be used as an antioxidant.
The pH of the extracted solution was 6.6±0.1 after desalting by ultrafiltration. Ko et al. (2011) reported that 10% (NH4)2SO4 used to extract phosvitin from egg yolk granules after removing lipids with ethanol had the highest recovery rate at pH 4. Thus, the pH of the extracted solution after desalting was adjusted to 4, 5, and 6, centrifuged at 4,070×g for 30 min, and the supernatants were lyophilized. However the extracted phosvitin recovery rates were not significantly improved by this pH adjustment. Additionally, the iron-binding ability of the separated phosvitin at pH 4 was significantly lower than that at pH 6.6±0.1 (data not shown). This result confirmed that the iron-binding ability of phosvitin was the highest in neutral pH (6.5) conditions (Castellani et al., 2004).
CONCLUSION
Our results suggest that separation of phosvitin from egg yolk can be done without using organic solvents, but that a thermal treatment (85 to 90°C) is needed to remove impurities from the 10% NaCl or (NH4)2SO4 extracts. Although the recovery rate of phosvitin from yolk granules was lower than that from other methods, this method was relatively simple, cost-effective, environmentally friendly, and suited for large-scale production. The extracted phosvitin was confirmed to have very high ferrous and ferric iron-binding ability, which is important for its applications in food and other industries.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







