





INTRODUCTION
The rumen epithelium plays a vital role in facilitating nutrient absorption and metabolization in ruminants, with its health status directly impacting the health of ruminants. The rumen epithelial barrier is composed of a multi-cellular structure and intercellular linking complex, which effectively prevents the translocation of lipopolysaccharide (LPS) into the bloodstream. A high-concentrate diet can induce ruminal acidosis, leading to the release of LPS, which are cell wall components of gram-negative bacteria in the rumen. A high concentration of LPS in the rumen can impair the structural integrity of the rumen epithelium and ruminal epithelial barrier function, leading to translocation into bloodstream and triggering an inflammatory response in ruminant organisms [1]. Cells possess an autonomous and orderly self-control metabolic system, which can yield a plethora of experimental materials. Cell culture enables direct observation of the morphological structure and vital activities of cells, making it a widely utilized tool in various disciplines such as cytology, genetics, immunology, experimental medicine and oncology. Cell culture offers a cost-effective alternative to the study of large animals (dairy cow, sheep, etc). Previous study found cultured ruminal epithelial cells respond to LPS stimulation by increasing transcription of pro-inflammatory genes and this transcriptional response was influenced by the dose, duration, and frequency of LPS exposure [2]. This establishes a basis for investigating the impact of acidosis on rumen epithelial function at a mechanistic level.
Antibiotics, such as monensin, play an important role in preventing rumen acidosis in ruminants. However, the large amounts of antibiotics used in ruminant fodder generate bacterial drug resistance and drug residues in animal products that can present a serious threat to human health. Therefore, there is an urgent need to identify substitutions for the current antibiotics. A moderate level of flavonoids increased growth performance and decreased the risk of ruminal acidosis in cattle on high-concentrate diet, which have the similar effects of monensin [3]. Hence, antibiotics may be replaced by flavonoids in ruminant production. Rutin, or rutoside, quercetin-3-O-rutinoside, or vitamin P, is a natural flavonoid glycoside that is a secondary metabolite in plants, which has antioxidant, anti-hypertension, anti-diabetes, anti-inflammation, and cardioprotective functions. Previously, we reported that rutin promoted the growth performance of sheep by enhancing antioxidant capacity and rumen fermentation and regulating hormone secretion, and improved meat quality of goats [4ŌĆō6]. These studies indicate that rutin might be used in ruminant production as a feed additive. Its absorption and metabolism is the premise of the biological functions of rutin. Rutin is stable in gastric and small intestinal fluid for 2 to 4 h and can be absorbed by the gastrointestinal tract. Guo [7] reported that the peak plasma concentration of rutin and quercetin in dairy cows was at 1 h and 8 h after intake, respectively. Previous studies found rutin could relieve ethanol-induced gastric lesions and alleviate ischemia-reperfusion-induced injury of rat small intestine, respectively [8,9]. In addition, rutin has higher bioavailability in ruminants compared to monogastric species. Hence, rutin can be absorbed and metabolized in the gastrointestinal tract, then play a role in protecting the rumen and intestine of ruminants.
According to the study of Shi et al [10], an increase in LPS concentration and incubation time resulted in a significant reduction of the relative growth rate of cell. LPS can decrease the activities of antioxidant enzymes glutathione peroxidase (GSH-PX), catalase (CAT) and superoxide dismutase (SOD), elevate the malondialdehyde (MDA) content, concentrations of pro-inflammatory cytokines interleukin-1 (IL-1) and IL-6 as well as nitric oxide (NO), and induce the expression of iNOS in bovine mammary epithelial cells. In addition, LPS-induced primary goat mammary epithelial cells for 3 h led to an increase in expression of C-C motif chemokine ligand 2 (CCL 2), C-X-C motif chemokine ligand 6 (CXCL 6), IL-6, CXCL 8, prostaglandin-endoperoxide synthase 2 (PTGS 2), interferon induced protein with tetratricopeptide repeats 3 (IFIT 3), myeloid differentiation primary response gene 88 (MyD88), nuclear factor of kappa light polypeptide gene enhancer in B-cells 1 (NFKB1) and toll-like receptor 4 (TLR 4) [11]. Our previous study revealed a significant decrease in the activities of GSH-PX, CAT, and SOD, as well as a significant elevation in the relative mRNA transcript levels of cytokines (CCL 5, CXCL 6, CXCL 8, IL-6, tumor necrosis factor-╬▒ [TNF-╬▒], and IL-1╬▓) and regulatory factors (TLR 2, nuclear factor ╬║B [NF-╬║B], and MyD88) in LPS-induced goat rumen epithelial cells (GRECs) [12]. These studies have demonstrated that LPS can elicit proinflammatory effects in cells by inducing oxidative stress. Previous studies found rutin can attenuate LPS-induced inflammation in muscle cells and apoptosis in human bronchial epithelial cell line cells by inhibiting oxidative stress [13,14]. It showed that rutin has the role of protecting cells. Quercetin (aglycones of rutin) could alleviate LPS-induced GRECs damage by enhancing antioxidant capacity and inhibiting inflammatory response [12]. At present, there has been no report on the protective effect of rutin against LPS-induced injury in GRECs. To verify whether rutin can protect GRECs like as quercetin, we examined its effects on the oxidation resistance, anti-inflammation, and barrier function of LPS-induced GRECs. This study was aimed at providing a theoretical basis for future studies on rutin protection of the ruminal epithelium in vivo.
MATERIALS AND METHODS
This experiment was performed according to the guidelines approved by the Animal Ethics Committee of Animal Husbandry and Veterinary, Jiangxi Academy of Agricultural Sciences (2010-JAAS-XM-01).
Cell culture and experimental design
The GRECs obtained from Boer goat ruminal epithelium were from the Institute of Animal Culture Collection and Application (IACCA), Yangzhou University. The GRECs (1├Ś105 cells/mL) were suspended with basal medium (DulbeccoŌĆÖs modified EagleŌĆÖs medium [DMEM]/F12 medium), inoculated into culture plates, and cultured in an incubator at 37┬░C with 5% CO2 saturated humidity until they had completely adhered to the plates. The GRECs were subcultured until the third passage. When the adherence of GRECs reached 80% to 90%, the GRECs were digested with trypsin and washed twice with phosphate-buffered saline (PBS). Then, the GRECs were divided into three groups and cultured in basal medium without rutin or LPS (control group, Con) or basal medium containing 1 ╬╝g/mL LPS (L), or 1 ╬╝g/mL LPS and 20 ╬╝g/mL rutin (L+R). Six replicates were performed for each group. The basal medium was supplemented with rutin (Ōēź98% purity, Yuanye Bio-Technology, Shanghai, China) dissolved with dimethyl sulfoxide (DMSO, Cell level; Sigma-Aldrich, St. Louis, MO, USA). All experimental groups received 0.1% DMSO. After 3-h culture, the GRECs were harvested to detect the relevant parameters.
Cell proliferative activity
GREC proliferative activity was detected with Cell Counting Kit-8 (Beyotime Biotechnology, Shanghai, China). The GRECs (1├Ś104 cells per well) were inoculated into 96-well plates and incubated at 37┬░C with 5% CO2 saturated humidity for 3 h, then incubated for 3 h with 10 ╬╝g/mL CCK-8 solution. Then, the absorbance of the GRECs was assayed at a wavelength of 450 nm using a microplate spectrophotometer (Multiskan Spectrum; Thermo Fisher Scientific, Waltham, MA, USA). The cell viability was calculated as follows:
where At, Ac, and Ab are the absorbances of the treatment groups, control group, and blank group (containing only basal medium), respectively.
Determination of transepithelial electrical resistance
The GRECs (1├Ś105 cells per cm2) were seeded into a transwell chamber (Costar, Coring Inc, New York, NY, USA) with 4.5 ╬╝m pores fit-ted into a 6-well plate. The other one transwell was used as a blank, which did not inoculate the GRECs. Transepithelial electrical resistance (TEER) of all treatment groups was measured by an epithelial volt-ohm meter with a chopstick electrode (Milli-cell ERS-2; EMD Millipore, Billerica, MA, USA) after finishing incubation of GRECs. The electrode was inserted the basolateral chamber and the apical chamber, respectively. Each monolayer was detected three times, and paid attention to preventing electrode contact with the monolayer. The mean resistance of a blank was deducted from all treatment groups. Finally, the unit area resistance was calculated by dividing resistance values by effective membrane area (4.5 cm2).
Apoptosis assay
GREC apoptosis was assayed by an annexin VŌĆōpropidium iodide (PI) apoptosis detection kit (Absin, Shanghai, China). The GRECs (1├Ś105 cells/mL) were seeded in 6-well plates, cultured in the treatment media for 3 h, detached with trypsin, then collected after washing twice with cold PBS. The GRECs were suspended with tag solution containing annexin V and PI, then incubated in the dark for 15 min at room temperature. The subsequent steps were performed according to the kit instructions. Finally, the cell fluorescence was detected using a flow cytometer (CytoFLEX; Beckman Coulter, Brea, CA, USA). The wavelength of exciting light is 488 nm. Fluorescein isothiocyanate fluorescence was detected by a passband filter with a wavelength of 515 nm and PI was detected by filter with wavelength greater than 560 nm. The left bottom of quadrant shows living cells, the upper right of quadrant shows necrosis cells, and the right bottom of quadrant shows apoptosis cells on the scatter plot of the bivariable flow cytometry.
Anti-oxidative parameters
Cell suspensions (2 mL) containing 1├Ś105 GRECs/mL were inoculated into 12-well plates. The basal medium was replaced by the treatment medium after cell attachment. The GRECs were incubated for 3 h in an incubator and then collected for detecting SOD, GSH-PX, CAT, and lactate dehydrogenase (LDH) activity and MDA concentrations according to the kit instructions. The anti-oxidative parameter detection kits were all from the Nanjing Jiancheng Bioengineering Institute (Nanjing, China).
Reactive oxygen species (ROS) were detected as follows: GRECs cultured in treatment medium for 3 h were harvested and incubated for 1 h in basal medium containing 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA). Subsequently, the GRECs were trypsinized and washed twice with PBS before resuspension with 300 ╬╝L PBS. The fluorescence of the GRECs was measured by flow cytometry. The excited and emitted wavelength of flow cytometry is 488 nm and 525 nm, respectively.
Immunofluorescence assay of tight junction protein zonula occludin 1
After incubation in the treatment medium, the GRECs were fixed in 4% paraformaldehyde for 30 min, washed thrice with PBS, and sealed with 3% horse serum at room temperature for 30 min. After the horse serum had been removed, the GRECs were incubated with primary antibody against zonula occludin 1 (ZO-1) (Abcam, Shanghai, China) overnight in a refrigerator at 4┬░C, then washed thrice with PBS. Next, the GRECs were incubated for 1 h in the dark at room temperature with the secondary antibody (Abcam, China) containing green fluorescence, then incubated in DAPI staining solution (Solarbio, Beijing, China) at room temperature for 5 min. Finally, the cells were washed three times in PBS and observed and visualized using a fluorescence microscope (Olympus, Tokyo, Japan).
Real-time fluorescence quantitative polymerase chain reaction
Total RNA was extracted from the GRECs according to the instructions of an RNA extraction kit (Tiangen Biotech, Beijing, China). Reverse transcription was performed using a reverse transcription kit (Takara, Dalian, China). The reverse transcription was performed in a reaction mixture with a total volume of 20 ╬╝L containing 10 ╬╝L master mix, 1 ╬╝L PrimeScript RT Enzyme Mix I and RT Primer Mix, 4 ╬╝L 5├Ś PrimeScript Buffer 2 and RNase-free dH2O. The reverse transcription reaction condition was 37┬░C for 15 min and 85┬░C for 5 s.
The real-time fluorescence quantitative polymerase chain reaction (qRT-PCR) was performed using a SYBR Premix Ex Taq II kit (Takara, China). The qRT-PCR primers were synthesized by Invitrogen (Shanghai, China); Table 1 lists their sequences. The primer sequences of genes excepting Claudin-1 and Occludin in Table 1 were from the thesis of Hu et al [15]. The qRT-PCR was performed in a total volume of 20 ╬╝L containing Premix Ex Taq II (10 ╬╝L), forward primer (0.8 ╬╝L), reverse primer (0.8 ╬╝L), complementary DNA (cDNA) template (2 ╬╝L), and PCR-grade water (6.4 ╬╝L). The qRT-PCR was performed on a LightCycler 96 real-time PCR instrument (Roche, Basel, Switzerland) under the following conditions: initial denaturation at 95┬░C for 30 s, followed by 40 cycles of PCR at 95┬░C for 5 s and 60┬░C for 30 s. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the reference gene for calculating the relative expression of the target genes with the comparative threshold cycle (2ŌłÆ╬ö╬öCt) method.
Determination of protein content by enzyme-linked immunosorbent assay
First, the supernatant of the GRECs was collected and used for measuring the protein contents of cytokines (IL-1╬▓, IL-6, and TNF-╬▒) and chemokines (CXCL6 and CXCL8). Subsequently, the GRECs were collected for extracting protein. The proteins extracted from GRECs were used for detecting the protein contents of key molecules in TLR/NF-╬║B signal pathway, such as toll-like receptor 2 (TLR2), TLR4, NF-╬║B, and inhibitor of nuclear factor kappa-B alpha (I╬║B╬▒). The method of protein extraction was as follow. The GRECs were added 1 mL extracting solution and crushed by ultrasonic wave (ice bath, 200 W, ultrasonic for 3 s, 10 s for interval, repeat 30 times). Then, the supernatant was collected by centrifuging at 4┬░C, 12,000 rpm for 10 min after finishing the ultra sonication. The protein content of supernatant was detected according to the instruction of BCA protein assay kit (Solarbio, China). Finally, the contents of those proteins were detected according to the manufacturerŌĆÖs instructions of the enzyme-linked immunosorbent assay (ELISA) kits (FANKEW; Shanghai, China). The ELISA kits of IL-1╬▓ (F6400-B; FANKEW, China), IL-6 (F3885-B; FANKEW, China), TNF-╬▒ (F3849-B; FANKEW, China), CXCL6 (F7100187-B; FANKEW, China), CXCL8 (F7100172-B; FANKEW, China), TLR2 (F7100169-B; FANKEW, China), TLR4 (F7100176-B; FANKEW, China), NF-╬║B (F7100181-B; FANKEW, China) and I╬║B╬▒ (F7100194-B; FANKEW, China) were bought from Shanghai Kexing Trading Co., Ltd (Shanghai, China).
Statistical analysis
Data were pretreated using Excel in WPS Office 2019 (Kingsoft, Beijing, China) and analyzed using one-way analysis of variance in SPSS 21.0 (IBM, Chicago, IL, USA). A Duncan multiple comparison test was performed and the data are reported as the means and standard errors of the mean (SEM). Statistically significant differences between treatment groups were determined at p<0.05, while extremely significant differences were observed at p<0.01. 0.05<p<0.10 was taken as an indication of tendency.
RESULTS
Effects of rutin on the cell activity of LPS-induced GRECs
Effects of different concentrations of rutin on the cell activity of GRECs are showed in Figure 1A. The cell activity of GRECs cultured in the medium supplemented 20 ╬╝g/mL of rutin was higher than that in other groups except the 80 ╬╝g/mL of rutin (p<0.05). Thus, we chose 20 ╬╝g/mL as the concentration of rutin in the following experiment. Effects of rutin on the cell activity of LPS-induced GRECs are exhibited in Figure 1B. The cell activity of GRECs was significantly reduced in the L group compared to the other groups (p<0.01) and was highest in the L+R group (p<0.01). Those results indicated that rutin had the function of improving cell viability of LPS-induced GRECs.
Effects of rutin on the permeability of LPS-induced GRECs
The results of the permeability of GRECs are displayed in Figure 2. Compared to the control group, the TEER of GRECs in the L group was significantly reduced (p<0.01). Meanwhile, the rutin can significantly enhance the TEER of GRECs induced by LPS (p<0.01). The results indicated that rutin can increase the TEER and decrease the the permeability of GRECs.
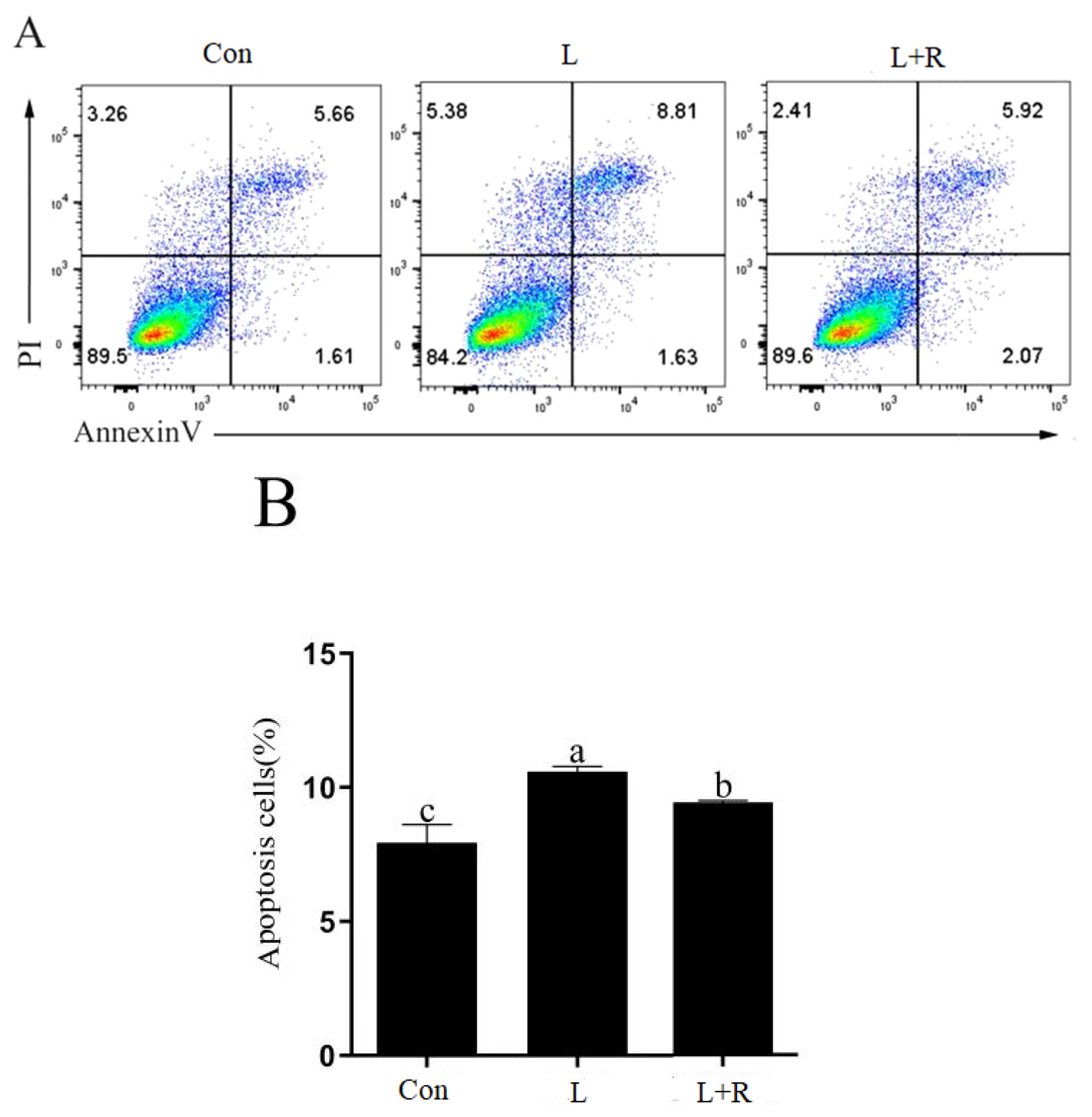
Effects of rutin on the apoptosis of LPS-induced GRECs
The apoptosis of GRECs in different treatment groups are showed in Figure 3A and 3B. Compared to the control, apoptosis was significantly increased in the L and L+R groups (p<0.05). Apoptosis in the L+R group was significantly lower than that in the L group (p<0.05). The results showed rutin could inhibit apoptosis of GRECs induced by LPS.
Effects of rutin on the antioxidative parameters of LPS-induced GRECs
The results for the GREC antioxidative parameters are depicted in Table 2. Compared with the control, SOD, GSH-PX, and CAT activity in the L group was significantly decreased, but LDH activity and ROS and MDA levels were significantly increased (p<0.01). Compared to the L group, SOD, GSH-PX, and CAT activity in the L+R group was significantly enhanced but LDH activity and ROS and MDA levels were significantly reduced (p<0.01). The results suggested that rutin supplementation enhances the antioxidant capacity of LPS-induced GRECs.
Effects of rutin on relative expression of cytokine genes in LPS-induced GRECs
The mRNA relative expression of cytokine genes in different treatment groups are displayed in Table 3. Compared to the control, the mRNA relative expression of IL-6 (p<0.05), TNF-╬▒ (p<0.01), and IL-1╬▓ (p<0.01) was enhanced significantly in the L group. Rutin significantly reduced the mRNA relative expression of TNF-╬▒ in the LPS-induced GRECs (p<0.01), whereas it had no significant effect on IL-6 and IL-1╬▓ mRNA relative expression (p>0.05). Compared to the control group, CCL5, CXCL6, and CXCL8 mRNA relative expression was significantly increased in the L and L+R groups (p<0.01). CXCL6 and CXCL8 mRNA relative expression was significantly lower in the L+R group than in the L group (p<0.01). The results indicated that rutin might inhibit the inflammatory reaction of GRECs induced by LPS.
Effects of rutin on the relative expression of regulatory factor genes in LPS-induced GRECs
The mRNA relative expression of regulatory factor genes in GRECs from different treatment groups are displayed in Table 4. Compared to the control, TLR2 (p<0.01), TLR4 (p<0.05), and NF-╬║B (p<0.01) mRNA relative expression was significantly increased in the L. Compared to the L group, TLR2 (p<0.01), TLR4 (p<0.05), NF-╬║B (p<0.01) and interferon (IFN) regulatory factor 3 (IRF3) (p<0.05) mRNA relative expression was significantly decreased in the L+R group. MyD88 mRNA relative expression in the L+R group was significantly higher than that in the the control (p<0.05), whereas interferon induced protein with tetratricopeptide repeats 3 (IFIT3) and toll-interacting protein (TOLLIP) mRNA relative expression was not significantly different between the groups. The results showed rutin might suppress the inflammatory reaction in LPS-induced GRECs by regulating the expression of TLRs and NF-╬║B.
Effects of rutin on protein contents of cytokines in LPS-induced GRECs
The protein contents of cytokines in cells are showed in Table 5. Compared to the control, the protein contents of TLR4, NF-╬║B, IL-1╬▓, and CXCL8 in the L group was significantly increased (p<0.05), whereas the protein content of TLR2 had no significant difference in all treatment groups (p<0.05). Compared to the L group, the protein contents of TLR4, IL-6, TNF-╬▒, and CXCL8 in the L+R group was significantly decreased (p<0.05). The protein contents of NF-╬║B and CXCL6 had a decrease tendency in the L+R group compared with the L group (p = 0.06). Compared to the L group, the protein content of I╬║B╬▒ in the L+R group had an increasing tendency (p = 0.08). The results indicated that rutin might affect the inflammatory reaction in LPS-induced GRECs by regulating secretion of cytokines protein.
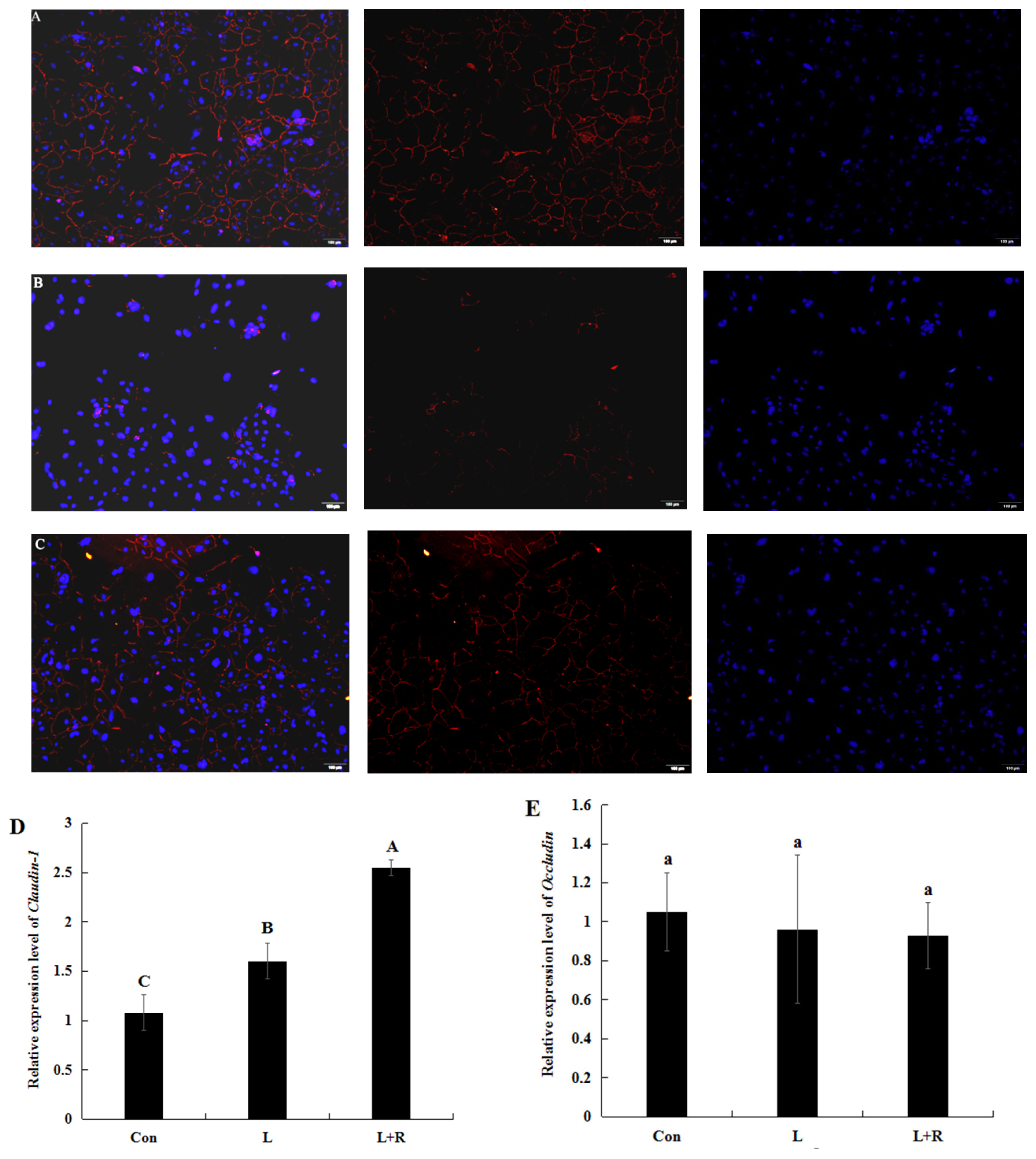
Effects of rutin on tight junction protein expression in LPS-induced GRECs
The expression level of the tight junction (TJ) protein (ZO-1, Claudin-1, and Occludin) in the GRECs from different treatment groups are depicted in Figure 4A to 4E. The edge of the LPS-induced GRECs exhibited slight red spotty staining and fewer continuous grid structures (Figure 4B), whereas the edges of rutin-supplemented GRECs demonstrated continuous grid structure (Figure 4C). This indicated that rutin enhanced ZO-1 expression levels in the LPS-induced GRECs. Claudin-1 (CLDN) mRNA relative expression in the L+R group was significantly higher than that in the L group (p< 0.01), while it was lowest in the control (Figure 4D). Occludin (OCLN) mRNA relative expression (Figure 4E) had no significant difference in all groups (p<0.05). It illustrated that rutin might improve the barrier function of LPS-induced GRECs in this study.
DISCUSSION
Shi et al [10] found that the relative growth rate of bovine mammary epithelial cells was significantly decreased with increasing the concentration of LPS and the incubation time. In addition, the cell viability of GRECs was reduced with increasing the LPS concentration in a certain range [16]. It showed LPS exerts a negative effect on cell proliferation. This study agreed with it. Previously, we found that quercetin supplementation could enhance the proliferation of LPS-induced GRECs [13]. The results of this study are consistent with it. However, according to the study of Kent-Dennis et al [2], LPS exposure did not negatively affect cell viability of bovine ruminal epithelial cells, which was different with us. The reason of it may be relation to different LPS concentration and treatment time. Apoptosis is a form of programmed cell death that plays a vital role in regulating cell development, cell aging, and the immune reactions of cells in tissues. Dai [17] reported that LPS induced by long-term high-concentrate diets induced rumen epithelial cell apoptosis in vivo and our results agree with this. The results of study by Paudel et al [14] showed rutin has the effects of potent anti-apoptotic activity against LPS induced damage. We also observed the similar results. In conclusion, rutin could promote the proliferation and inhibited the apoptosis of LPS-induced GRECs.
In general, the pro-oxidant and antioxidant systems in the body are in equilibrium. If a redox steady-state is imbalanced, it can lead to high free radical production. The large accumulation of free radical can induce unsaturated fatty acids to produce MDA in cells, and MDA promotes cell death by disintegrating cells. Antioxidative enzymes can scavenge free radicals and maintain a redox steady-state in the body. An increased production of ROS or a decreased defense capability of cellular antioxidant can cause oxidative stress. ROS produced in the process of oxidative metabolism could initiate the inflammatory process resulting in synthesis and secretion of proinflammatory cytokines. Furthermore, excessive ROS accumulation can cause irreversible damage to cells, which lead to cell death by the necrotic and apoptotic processes [18]. Previous studies showed that LPS can reduce the activity of antioxidative enzymes including SOD, CAT, and GSH-PX and enhance ROS and proinflammatory cytokine (IL-6, TNF-╬▒, and IL-1╬▓) production in cells [10,12]. The results of our study agreed with those studies. Our research findings indicate that LPS could trigger oxidative stress, leading to cell apoptosis. The chemical structure of rutin contains phenolic rings and free hydroxyl groups that can donate electrons to reactive free radicals to form more stable species and quench the free radical chain reaction [19]. Liu et al [13] and Paudel et al [14] found that LPS induced reactive ROS production of cells was significantly reduced by rutin. In addition, rutin could reverse decreased activities of antioxidative enzymes such as SOD, CAT, GSH-PX, and heme oxygenase-1 induced by LPS and improve the stressed condition by increasing the levels of GSH-PX, SOD, and decreasing MDA level [20,21]. Our study found the similar results. A cytosolic enzyme, LDH has been used as a sensitive index for assessing the degree of cell injury. Yang et al [22] reported that rutin reduced LDH levels in hypoxia/reoxygenation-induced H9c2 cells, and our results are consistent with that. Therefore, rutin could protect GRECs against damage by strongly scavenging free radical and inhibiting of lipid peroxidation.
Rutin plays a role in inhibiting the inflammatory response. According to the results of Su et al [23], rutin can decrease the protein and mRNA levels of TNF-╬▒, IL-1╬▓, and IL-6 in mammary tissues of mastitis model induced by LPS. Previous studies displayed rutin can suppress the production of TNF-╬▒ in LPS-induced human umbilical vein endothelial cells and decrease IL-6 and TNF-╬▒ gene expression in mouse muscle cells induced by LPS, which were similar with us [13,24]. Chemokines are a class of cytokines that regulate infiltrating cell migration and adhesion to an inflamed lesion. Based on the arrangement of the amino terminal cysteine, chemokines can be divided into four subgroups: C, CC, CXC, and CX3C. CCL5 facilitates the inflammatory responses and induces T cell subset adhesion and migration in immune responses. Known as granulocyte chemotactic protein 2, CXCL6 can be upregulated after stimulation by inflammatory factors. Proinflammatory cytokines, such as TNF-╬▒ and IL-1 can induce the expression of CXCL6. Also termed IL-8, CXCL8 is a proinflammatory chemokine rapidly induced by proinflammatory cytokines such as TNF-╬▒ and IL-1, bacterial or viral products, and cellular stress. The upregulation of CCL5, CXCL6, and CXCL8 in the LPS-induced GRECs is consistent with the results of Bulgari et al [11] and Hu et al [15]. Here, rutin reduced mRNA and protein levels of CXCL6 and CXCL8 in LPS-induced GRECs, which may be related to the reduced TNF-╬▒ levels. Therefore, rutin could attenuate LPS-induced inflammatory injury in GRECs by inhibiting the production of proinflammatory cytokines, such as TNF-╬▒.
Toll-like receptors (TLRs) are pattern recognition receptors (PRRs) that can recognize pathogen-associated molecular patterns (PAMPs) and activate signaling pathways dependent on the adaptors MyD88 or TRIF, and then activate NF-╬║B and induce the release of proinflammatory cytokines, chemokines, and type I interferon (IFN). IFN regulatory factor 3 (IRF3) is the most important transcription regulatory factor of type I IFN expression and is activated in the MyD88-independent signaling pathway by TLR4. Hu et al [15] found that LPS can upregulate expression of TLR2 and NF-╬║B in GRECs, but can not affect relative expression of MyD88, IRF3, IFIT3, and TOLLIP. In addition, LPS leads to increase in expression of TLR4 and NF-╬║B in goat mammary epithelial cells [11]. Our study found the similar results. According to the results of previous studies, rutin downregulated the expression levels of TLR4 and NF-╬║B, and inhibited the phosphorylation of NF-╬║B in LPS-induced cells and tissues [13,20,25]. Furthermore, rutin mitigated inflammatory reaction in cells and tissues by inhibiting the protein or mRNA expressions of TLR2, TLR4, and IRF3 [26], and our results are consistent with these findings. I╬║B╬▒ can inhibit the activation of NF-╬║B, and then inhibited the transcriptional activation of genes of proinflammatory factor. The results of our study showed rutin might inhibit the activation of NF-╬║B by increasing the level of I╬║B╬▒. Hence, rutin might reduce the expression of proinflammatory chemokines (TNF-╬▒) and chemokines (CXCL6, CXCL8) in LPS-induced GRECs by inhibiting the TLR/NF-╬║B signaling pathway. A type I IFN-stimulated gene, IFIT3 has antitumor activity. We demonstrated rutin did not affect IFIT3 expression. This indicated that rutin might not affect the IFN secretion in cells. TOLLIP can regulate the inflammatory reaction by suppressing proinflammatory cytokines and improving anti-inflammatory secretion in cells. However, TOLLIP expression has no significant difference in all treatment groups. Thus, rutin could not suppresses proinflammatory cytokines in GRECs by regulating TOLLIP expression.
TEER is an important indicator which evaluate the permeability and integrity of the epithelial barrier. The higher the value of TEER, the lower the permeability of epithelial barrier. Hence, rutin can reduce the permeability of GRECs according to the result of our study. TJs, such as the Claudin, Occludin, and ZO family members, are the most important structure of the rumen epithelium barrier, regulating nutrient absorption in the epithelium and preventing microbe and toxin entry from the rumen into blood. SARA can elevate levels of LPS, high acidity, and hyperosmolality and impair ruminal epithelial barrier function by decreasing the expression of TJ proteins (Claudin-1, Occludin, ZO-1) and increasing rumen epithelium permeability [27]. However, it has also been reported that an increase of SARA-induced LPS can decrease the mRNA and protein expression of ZO-1 and increase the mRNA and protein expression of Claudin-1 [28]. This study had similar results with them. Our results confirmed that SARA induced ruminal epithelial barrier injury possibly related to the LPS-decreased TJ proteins. Previous study found that autophagy can increase the protein expression and stability of Claudin 1. Furthermore, the increased level of Claudin 1 stimulated autophagy by decreasing the level of the autophagy substrate under autophagy-inducing conditions [29]. The increased expression of Claudin 1 in LPS-induced GRECs might be relation to autophagy of cells in this study. IL-1╬▓ can inhibite the activity of transcriptional promoters of Occludin protein, leading to decrease the mRNA expression of Occludin, and then destroy the effects of TJ. Compared to the control, the expression of Occludin was slightly decreased in LPS-induced GRECs, might be relation to an increased level of IL-1╬▓. Flavonoids, such as quercetin, morin, naringenin, enhance barrier functions of intestinal epithelial cells stimulated with high glucose by increasing Claudin-1, Occludin, and ZO-1 expression [30]. We also found rutin can enhance the expression levels of Claudin-1 and ZO-1 in LPS-induced GRECs. This indicated that rutin could relieve ruminal epithelial barrier function injury in GRECs induced by LPS. Therefore, rutin may protect the ruminal epithelial barrier against SARA, which nevertheless requires verification in the future.
CONCLUSION
LPS decreased the activity and increased apoptosis of GRECs and caused barrier injury in GRECs by promoting oxidation stress and the inflammatory response. Rutin alleviated the barrier injury of LPS-induced GRECs by enhancing oxidation resistance and anti-inflammation, which may be connected with inhibition of the TLR/NF-╬║B signaling pathway.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







