





INTRODUCTION
The poultry industry is an essential part of the agricultural sector. It plays a crucial role in providing a considerable source of animal protein, such as meat and eggs, to meet the growing demands of the global population [1]. In the poultry industry, the microbiome of poultry birds is critical in determining their health, growth, and productivity [2].
The microbiota is the community of microorganisms living in a particular environment, such as bacteria, fungi, and viruses. The connection between the human microbiome and health has been studied for over a century [3]. Recently, attention has been paid to the significance of the association between livestock and microbiota. Previous research has investigated fluctuations in the gut microbiota throughout lactation and weaning in pigs [4] and the rumen microbiota in sheep and its impact on feed efficiency [5]. The gut microbiota is the community of microorganisms living in the digestive tract of animals and is indispensable for efficient nutrient digestion and absorption. Furthermore, it plays a vital role in defending the host against pathogenic microorganisms [6]. The gut microbiota changes over time in animals [7].
This study selected laying hens (Hy-Line Brown), the most balanced brown egg layer worldwide. The importance of the gut microbiota in laying hens, as in other livestock, is gaining attention. A healthy microbiota in laying hens can reduce the risk of food-borne diseases [8]. Furthermore, a thorough understanding of gut microbiota changes over time is required to select the appropriate feed additive, such as probiotics and prebiotics [9], for each stage of chicken growth to achieve healthy eggs. However, there have been limited studies on the basic physiology of livestock, and few studies have investigated the intestinal microbiota in laying hens at different growth stages. We suggest that the gut microbiota of laying hens undergoes dynamic changes over time and can serve as an important indicator of the overall health and development of the gut microbiome. Therefore, changes in the gut microbiota of laying hens were monitored from birth to the pre-laying stage.
MATERIALS AND METHODS
Animal trial
Laying hens (Hy-Line Brown, n = 80) were obtained from poultry farms (Korea Poultry Co., Anseong, Korea) and raised on a farm in Chuncheon, Korea, following Korean animal welfare guidelines. Birds were fed a commercial diet suitable for their growth stages. Starter and well-textured mash diets were provided ad libitum on days 0 to 43 and 44 to 101 (Table 1), with free access to water. A microcontroller (NodeMCU) continuously monitored the temperature and humidity. All experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of Kangwon National University (KW-220425-5).
Sample collection
Twenty birds were selected based on body weights at each sampling point, spaced at equal intervals. For feces collection, the birds were isolated in a clean plastic pen floored with sterilized aluminum foil and weighed. Ten birds were selected from the twenty whose feces had been collected, spaced at equal intervals, and euthanized by CO2 asphyxiation to collect ileal contents at 10, 21, 58, and 101 days (Supplementary Table S1). In short, 1 g of feces and ileal contents were placed into 1.7 mL tubes using a sterilized tip. In total, 80 fecal and 39 ileal content samples were stored at −70°C, until DNA extraction.
DNA extraction and 16S rRNA sequencing
DNA was extracted from 250 mg feces and ileal contents using the NucleoSpin Soil kit (Macherey–Nagel, Düren, Germany). Briefly, each sample was homogenized using 0.6 to 0.8 mm ceramic beads in NucleoSpin bead tubes and a Taco Prep bead beater (GeneReseach Biotechnology Corp., Taichung, Taiwan). Subsequently, DNA was extracted following the manufacturer’s instructions. The extracted DNA was stored at −20°C until further analysis. The V4 region of the 16S rRNA gene was amplified using TaKaRa Ex-Taq polymerase (TaKaRa Bio, Shiga, Japan) and universal primers (forward: 5′-GGA CTACHVG GGTWTCTAAT-3′ and reverse: 5′-GTGCC AGCMGCCGC GGTA A-3′) with the following amplification conditions: 94°C for 3 min, followed by 30 cycles at 94°C for 45 s, 55°C for 1 min, 72°C for 1.5 min, and finally at 72°C for 10 min [10]. Amplicons were purified using a QIAquick polymerase chain reaction (PCR) purification kit and normalized to 50 ng per sample using a Spark 10 M Multimode microplate reader (Tecan Group AG, Männedorf, Switzerland). DNA library construction and sequencing were performed using the Illumina MiSeq platform (eGenome Inc., Seoul, Korea) to generate paired-end reads of 2×250 bp.
Microbiome analysis
Quantitative Insights into Microbial Ecology 2 (QIIME 2) v.2021.4 (https://qiime2.org) and the SILVA 16S rRNA gene reference database were used to analyze microbiome communities [11]. Individual primers and adapters were trimmed from raw sequencing reads using the QIIME2 plugins cutadapt and demux and demultiplexed using in-house Perl scripts [12]. Demultiplexed reads underwent quality trimming, filtering, and chimeric sequence removal using the denoise-paired option in the Divisive Amplicon Denosiong Algorithm (DADA) 2 plugin [13]. The DADA2 denoise-pair options were as follows: 8 base pairs from the left were trimmed and truncated at 180 bp. The generated phylogenetic tree was used to analyze the microbial diversity of the samples. Multiple alpha and beta diversity indices were generated from the phylogenetic tree using core-metrics-phylogenetic, alpha-group-significance, and beta-group-significance options in QIIME 2.
Alpha diversity (Shannon, Faith’s phylogenetic distance, and Pielou’s evenness) was used to evaluate species richness, evenness, and phylogenetic distance of the microbiota. Beta diversity was assessed using nonmetric multidimensional scaling (NMDS) using the Bray–Curtis dissimilarity matrix between samples, which was conducted using the vegan package in R. NMDS is an ordination technique used to visualize data patterns in N-dimensional spaces. Adonis statistical tests utilizing 999 permutations were employed to evaluate how innate factors affect the microbial community during the growth stages. The amplicon sequence variants generated by DADA2 were assigned to taxonomic classifications using the SILVA 132 16S rRNA classifier and pre-trained using the QIIME 2 fit-classifier-naïve Bayes option.
Metagenomics prediction
Metagenomics function was predicted using Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt) that, which utilizes 16S rRNA marker gene sequences and references to previously published complete genome sequences [14]. The 16S rRNA gene copy numbers in the BIOM files were normalized and adjusted, and the metagenomes were predicted using precomputed Kyoto encyclopedia of genes and genomes (KEGG) orthologs. The predicted metagenomes were then aggregated based on a specific hierarchy level in the KEGG pathway using its metadata. Principle component analysis (PCA) was performed using STAMP v.2.1.3.
Statistical analysis
Statistical analyses were performed using R v.4.1.3. One-way analysis of variance was used to compare microbial abundances between groups, followed by post-hoc Tukey’s honest significant difference test for pairwise multiple comparisons. Statistical significance was set at p<0.05. Pearson’s correlation coefficient (R) and p-values obtained from simple linear regression were used to evaluate the correlation between microbial abundance and body weights.
RESULTS
Animal growth
The body weights of chickens rapidly increased as they grew. The trend was similar to that of the standard body weight of Hy-line Brown chickens (Figure 1). The chicken body weights at different growth stages were 74.94±15.71, 174.31±22.65, 660.55±37.86, and 1358.15±133.99 g at 10, 21, 58, and 101 days, respectively.
Sequencing statistics of the gut microbiome
After sequencing the V4 region of 16S rRNA and performing quality control of the sequences, 3,082,756,756 sequences (mean = 25,906±22,690) were obtained. Each fecal and ileal content sample generated 15,020±6,543 and 48,236±27,359 average reads, respectively. The reads generated in fecal samples of different growth stages were 15,031±7,562 on day 10; 13,178±4,869 on day 21; 13,591±6,329 on day 58; and 18,278 ±6,336 on day 101. Similarly, for the ileal contents at different growth stages, the reads generated were 31,379±11,301 on day 10; 32,479±22,727 on day 21; 61,786±19,238 on day 58; and 65,722±33,101 on day 101.
Assessing the community of gut microbiota using 16S rRNA sequencing
The community diversity was first explored to investigate the microbiomes of developing laying hens. Microbial communities in the feces and ileum exhibited distinct patterns of richness and diversity over time. While the richness of the fecal microbiota significantly increased with age at 10 and 21 days and the late stages (58 and 101 days) (p<0.05; Figure 2A), the ileal samples showed a significant increase from the early (10 and 21 days) to late stages (p<0.05; Figure 2D). Furthermore, as measured by Faith’s phylogenetic diversity, microbial diversity of the ileal samples significantly increased with the progression of growth stages, whereas no significant change was observed in fecal microbial diversity (p<0.001; Figure 2B).
NMDS using the Bray–Curtis dissimilarity method was used to visually represent the changes in the composition of the gut microbiome, which changed with growth. The NMDS plot revealed that the fecal and ileal samples could be categorized into three discrete groups based on their microbial composition: 10 days, 21 days, and late stages (58 and 101 days). Adonis statistical analysis indicated a significant association between the growth stage of chickens and intestinal microbiota composition (feces: R2 = 0.24, p<0.001; ileal contents: R2 = 0.23, p<0.001; Figure 2C and 2F).
Relative abundance of gut microbiota at different growth stages
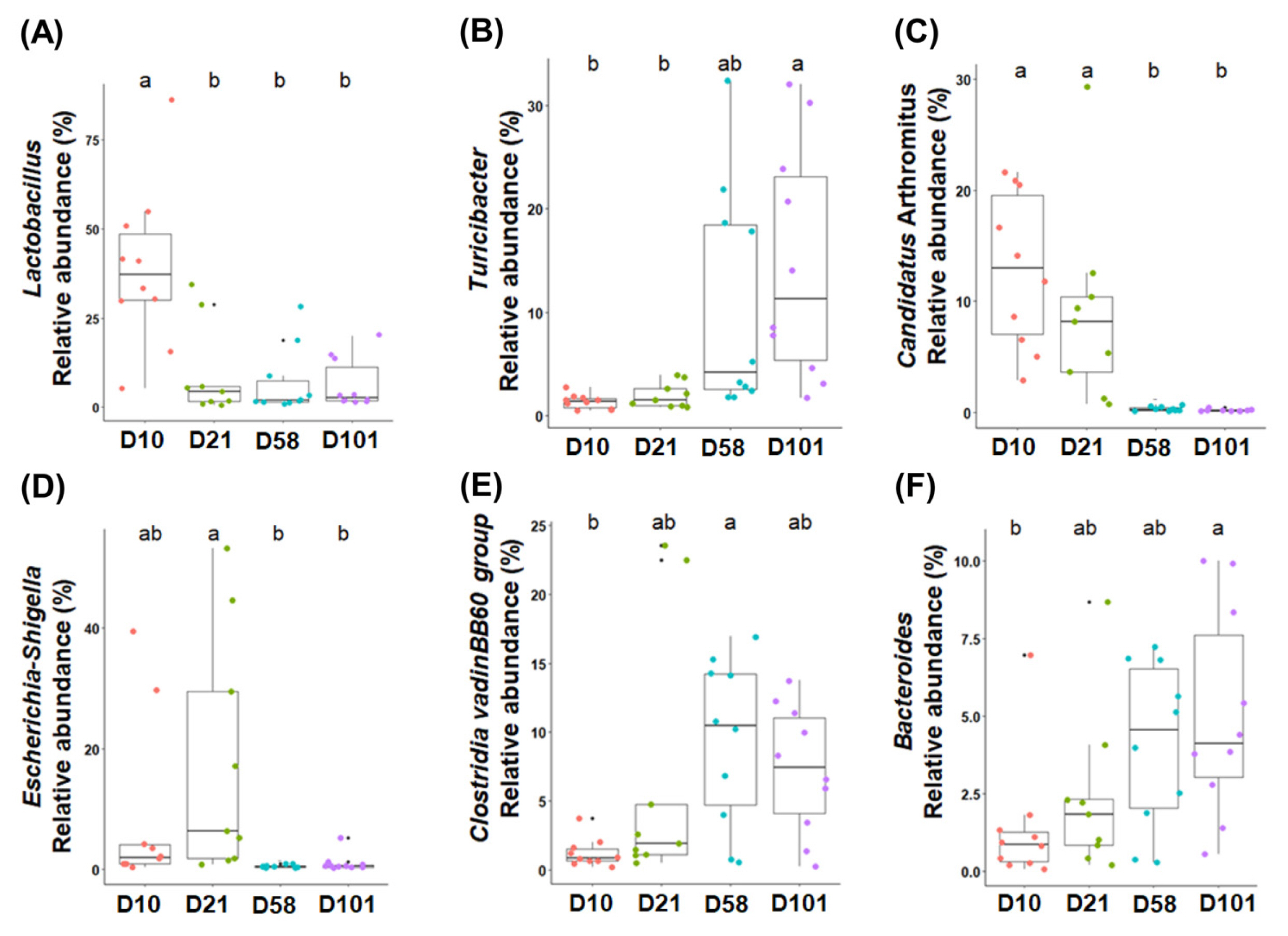
Differential analysis was performed to characterize the microbiota in detail to evaluate the relative abundances of all phyla and genera (Tables 2 and 3). The three major phyla in feces and ileal contents were Firmicutes, Proteobacteria, and Bacteroidota. Firmicutes had the highest relative abundance in both groups, and that of Proteobacteria significantly increased with growth and peaked at day 21. Although no significant difference in the abundance of Bacteroidota was observed in fecal samples, it significantly increased in the ileal samples. The major microorganisms identified in the feces at the genus level were Romboutsia, Lactobacillus, Streptococcus, Clostridium sensu stricto I, Escherichia-Shigella, and Turicibacter (Figure 3). The relative abundance of Romboutsia gradually increased with growth and peaked at day 58, whereas that of Lactobacillus was highest on day 10 and decreased with the growth of the chicken (Figure 3A and 3B). Streptococcus was most prevalent at day 21, whereas Clostridium sensu stricto I showed no significant differences. The abundance of Escherichia-Shigella was highest in the early stages and gradually decreased, whereas that of Turicibacter showed the opposite correlation. The abundances of Lactobacillus, Turicibacter, and Escherichia-Shigella in the ileal contents showed similar trends to those in the feces (Figure 4B, 4C, and 4D). The abundance of Clostridia vadin BB60 group was highest at day 58, and that of Candidatus Arthromitus decreased with growth; however, Bacteroides showed an opposite correlation.
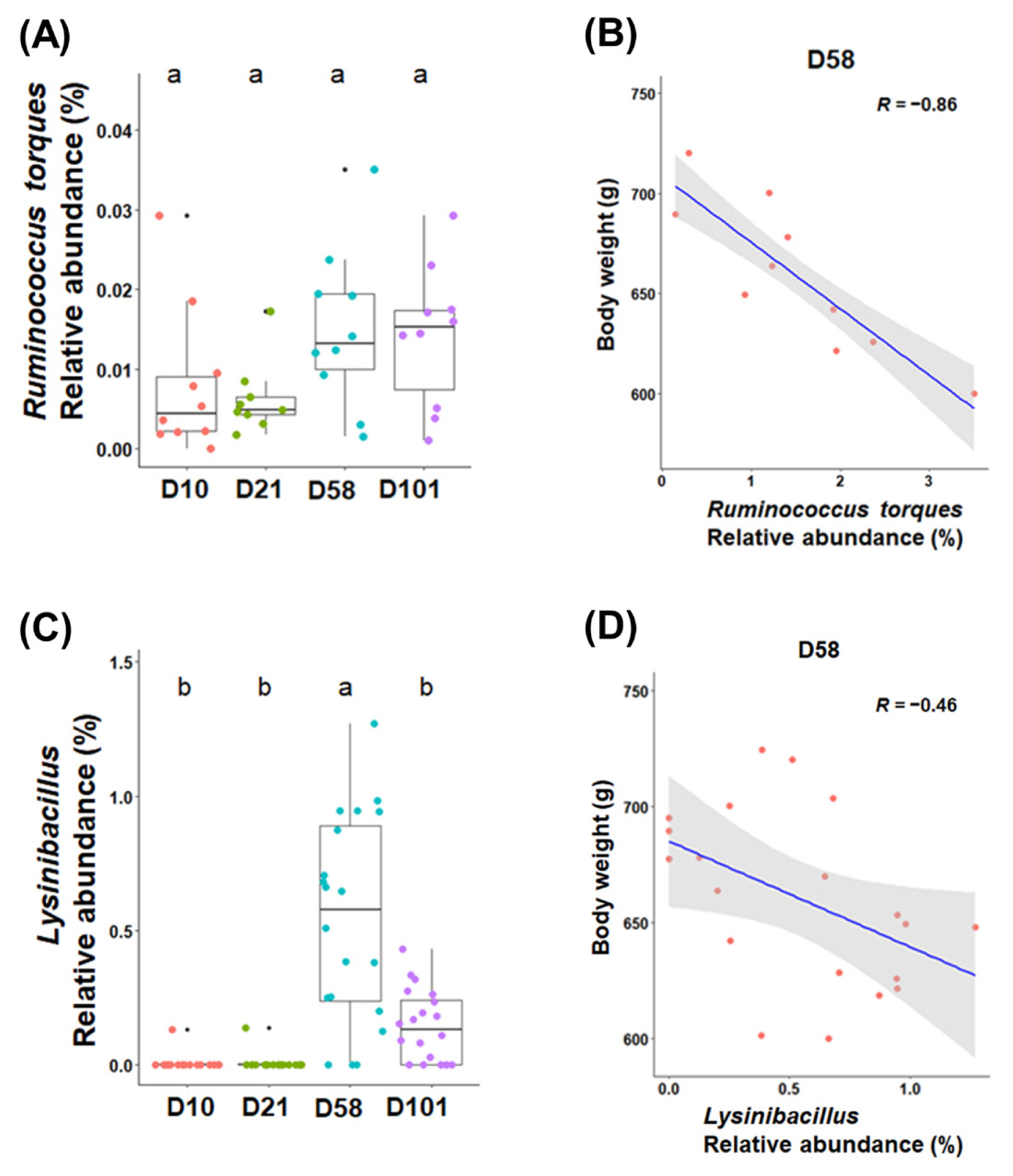
Linear regression analysis was conducted to identify the bacterial taxa correlated with body weight in different sampling sections of chickens. For the correlation between genera and body weight, in particular, the abundances of Ruminococcus torques (R = −0.86, p<0.001) and Lysinibacillus (R = −0.46, p<0.05) were negatively correlated with body weight (Figure 5).
Metagenomics pathways
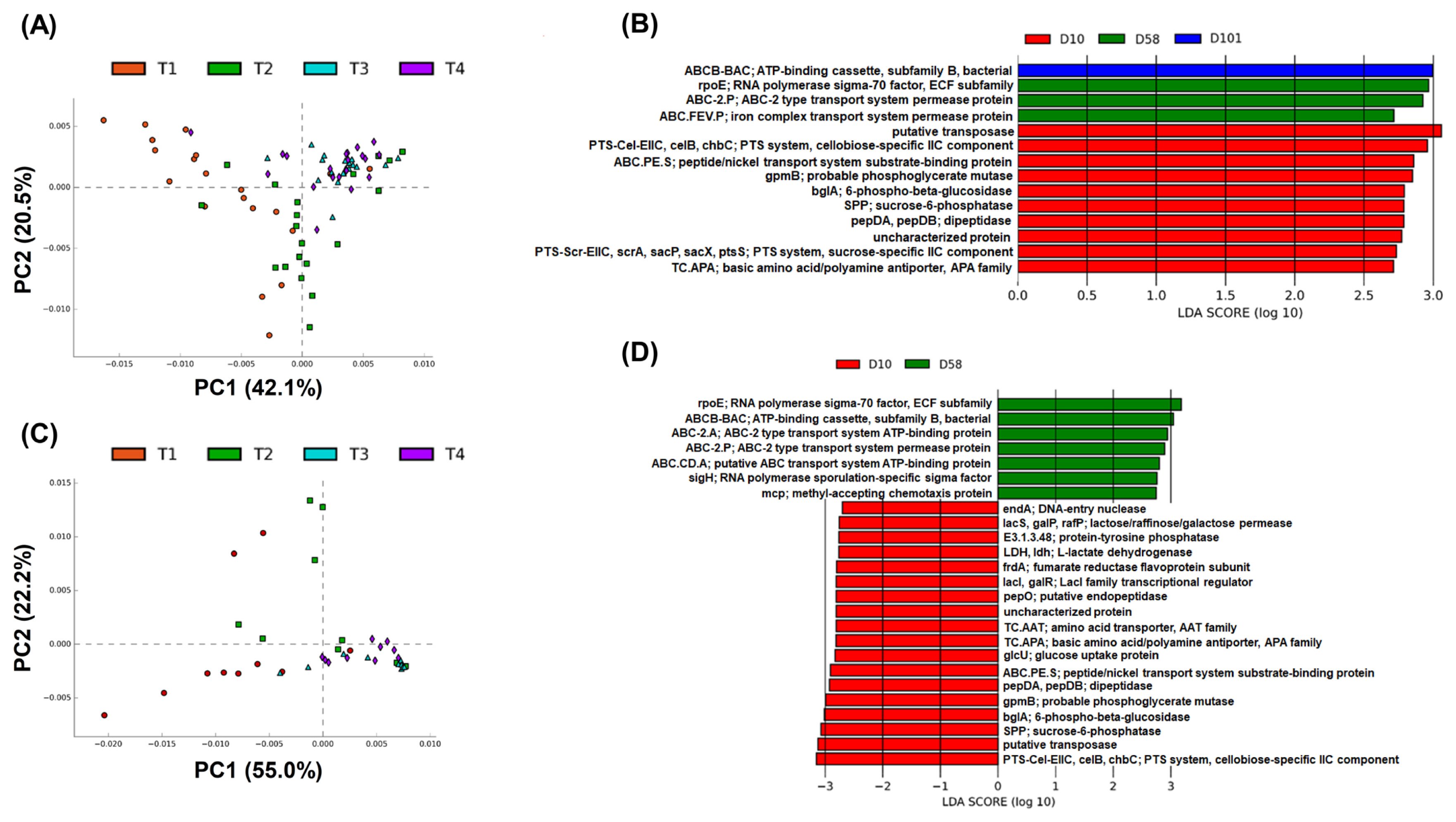
The KEGG pathways were used to compare the functions of gut microbiota at different growth stages. A total of 6,665 and 6,510 KEGG pathways were identified in the feces and ileal contents, respectively. PCA was conducted at level 4 of the KEGG pathways to observe the sample distribution pattern. For each group of samples, the resulting PCA plot revealed three distinct groups in which the samples were clustered (Figure 6A and B).
Next, linear discriminant analysis effect size (LEfSe; linear discriminant analysis [LDA] score >2.7 and p<0.05) was performed. Fourteen KEGG pathways were identified in the feces (Figure 6B). The pathways related to carbohydrate and energy metabolisms and “putative transposase” had significantly high scores on day 10, while those related to “RNA polymerase sigma-70 factor, ECF subfamily,” “ABC-2 type transport system permease protein” and “iron complex transport system permease protein” had significantly high scores on day 58. The pathways related to “ATP-binding cassette, subfamily B, bacterial” had high scores on day 101. Twenty-five KEGG pathways were identified in the ileal contents (Figure 6D). The pathway related to “PTS-Cel-EIIC, celB, chbC; PTS system, cellobiose-specific IIC component” had a significantly low score on day 10; “putative transposase,” “SPP; sucrose-6-phosphatase,” and “bglA; 6-phospho-beta-glucosidase” had LDA scores <3.0. Several nucleotide, carbohydrate, pyruvate, amino acid, and propanoate metabolic pathways showed significantly low scores on day 10. On day 58, “rpoE; RNA polymerase sigma-70 factor, ECF subfamily” and “ABCB-BAC; ATP-binding cassette, subfamily B, bacterial” had LDA scores >3.0. “ABC-2. A; ABC-2 type transport system ATP-binding protein,” “ABC-2. P; ABC-2 type transport system permease protein,” and “ABC.CD.A; putative ABC transport system ATP-binding protein” were related to signaling and cellular processes; “sigH; RNA polymerase sporulation-specific sigma factor” and “mcp; methyl-accepting chemotaxis protein” were related to genetics and signal transduction, respectively.
DISCUSSION
A longitudinal study was conducted on the gut microbiota of laying hens across four growth stages in two sampling sites. Longitudinal study refers to a study that tracks or observes the same subject over a long period. Previous studies have analyzed the gut microbiota of laying hens in the early, middle, and late laying stages [15] and broilers at different growth stages [16]. Nutritional responses, immune system interactions, and pathogen infection have been associated with the gut microbiota of laying hens [17]. Laying hens produce eggs after three months, meaning their breeding period is longer than that of broilers. Analyzing the gut microbiota based on the growth stage is crucial for stable breeding. However, there have been limited gut microbiota studies during detailed growth stages before laying in Hy-line hens. This study was conducted because investigating the gut microbiota before laying rather than after laying was considered necessary.
Alpha diversity was investigated to explore the gut micro biota composition in laying hens. Previous studies reported that microbial diversity increased with growth in broilers [18], laying hens [19], and mice [20]. Similarly, the microbial diversity and richness at 101 days were higher than in other growth stages, regardless of sampling sections. The relationship between growth and microbial diversity is affected by various factors, such as age, feed change, and breeding environment [21,22]. Likewise, microbial diversity significantly increased after feed change, implying that growth and feed change positively correlate with microbial diversity. However, further research is required to determine how feed relates to microbial composition.
NMDS analysis was conducted using Bray–Curtis dissim ilarity to validate the changes in the intestinal microbiota during growth. Samples collected on days 10, 21, 58, and 101 were grouped into four distinct clusters representing different age groups, with early-stage clusters comprising samples from days 10 and 21. In contrast, late-stage clusters comprised samples from days 58 and 101. Late-stage clusters differed from early-stage clusters in terms of age and diet composition. Similar alterations in the intestinal microbiota owing to age and diet have previously been observed [7]. For this study, two diets were administered based on the growth stages. Chicks were fed a starter feed on days 10 and 21, and a well-textured feed with modified nutrient content was provided on days 58 and 101 during preparation for laying. The starter feed comprised large particles with high crude protein and fat contents. In contrast, the well-textured mash diet contained crude fiber and ash at elevated levels in the form of pellets (Table 1). Providing appropriate feed according to different growth stages is crucial in the poultry industry because it affects nutrient availability, egg production, and overall productivity.
Changes in the microbiota of feces and ileal contents de pend on age and diet. The abundance of Lactobacillus was highest at day 10 in all groups and gradually decreased as the chickens grew. Broilers fed starter feed showed a similar trend [23]. Lactobacillus produces lactic acid in the gut, lowering pH and reducing the abundance of pathogens [24]. A high abundance of Lactobacillus in chicks positively affects intestinal health, reducing intestinal permeability and improving gut health [25]. Therefore, Lactobacillus in the early stages of chicken growth may positively affect health. Romboutsia was predominant in the feces and increased with age, reaching its highest relative abundance at day 58. Romboutsia is associated with feed utilization and efficiency in broilers owing to its carbohydrate utilization and fermentation of single amino acids [26]. Escherichia-Shigella and Streptococcus are partially pathogenic; they showed the highest relative abundances at day 21, which decreased as the chickens grew. Salmonella is affected by chicken age and immune system [27]. Likewise, both pathogens are thought to be affected by these factors. The abundance of Clostridium sensu stricto 1 in feces showed no significant differences between growth stages. This genus includes Clostridium perfringens and other pathogenic Clostridium species [28]. Candidatus Arthromitus is found in the terminal ileum of animals and has unique immunomodulatory properties [29]. The abundance of Candidatus Arthromitus decreases as chickens grow, and similar results have been observed in poultry [30]. This potentially occurs because immunity was increased during growth. Turicibacter has anti-obesity properties, reduces metabolic stress, and inhibits inflammatory reactions in rats [31]. Feeding well-textured feed in the late stages showed an increase in the body weight of chickens compared to that in the early stages. The study examined the relationship between the microbiota and body weight. R. torques exhibited a negative correlation with body weights in fecal samples. This microbiome belongs to the Clostridium coccoides (XIVa) group in humans and degrades gastrointestinal mucin [32]. While factors such as feed and environment affect the body weight of chickens, our findings highlight the significant influence of microbiota in this regard [33].
The PCA results of metabolic KEGG pathways were clus tered into three groups regardless of sampling sites: day 10, day 21, and late stage (days 58 and 101). LEfSe analysis revealed that the pathways associated with membrane transport, carbohydrate metabolism, and energy metabolism had high scores in the feces. These pathways are essential for bacterial growth to survive in the gut. “PTS-Cel-EIIC, celB, chbC; PTS system, cellobiose-specific IIC component” scored lowest at 10 days in the ileal contents. This is a major active-transport system for carbohydrates, which catalyzes the phosphorylation of incoming sugar substrates concomitant with their translocation across the cell membrane. Phosphorylation is an essential factor for bacterial growth and is regulated by infection in chickens [34]. We hypothesize that this microbial pathway was developed because chicks are vulnerable to infection. However, further studies are required to uncover the exact differences in metabolisms.
CONCLUSION
The four growth stages were determined to contribute to gut microbiota changes in laying hens. Similar to previous studies, the results showed that various factors, such as growth stage and feed, are crucial. Among them, it is essential to effectively manage pathogens and beneficial bacteria in the early growth stage because pathogen richness in the early growth stage may lead to a decrease in immunity. The results also suggest that there may be interactions between microbiota and feed. Our results can enhance the understanding of microbiology in the poultry industry and will be valuable data for applied research. The changes in the gut microbiota after laying would be an interesting topic for future research.



 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print







