





INTRODUCTION
Pseudomonas aeruginosa is a gram-negative bacterium that is widely known to account for up to 10% of all human and animal infections and is one of the main causes of numerous diseases such as hemorrhagic pneumonia, mastitis, urinary tract infections, otitis, and endometritis in domestic animals, causing tremendous damage to animal husbandry [1–3]. Not only does P. aeruginosa causes respiratory diseases in animals such as chickens and cattle, but it also causes mastitis in Holstein cows, and according to Banerjee et al [4], mastitis is a known cause of the loss of milk production, which makes it a major problem in the industry [4–7]. Infection with P. aeruginosa is stimulated by the microbial molecules called pathogen-associated molecular patterns (PAMPs) [8], and the bacterial flagellin is a well-known PAMP, shown to upregulate pro-inflammatory mediators [9,10]. Also, as it is widely known that the activity and properties of certain proteins vary greatly depending on the phosphorylation of corresponding protein [11], dephosphorylating bacterial flagellin might lead to the regulation of inflammation in flagellin-mediated infections. Structurally, P. aeruginosa possesses an unusual surface filament-like flagellum made up of post-translationally phosphorylated flagellin protein [12].
Sweet potato purple acid phosphatase (PAP) is a binuclear metal-containing phosphatase, meaning that it has enzymatically active binuclear metal sites (Fe-Mn center) [13]. The structure of sweet potato PAP is very similar to that of enzyme called phosphoprotein phosphatases (PPPs), binuclear metallohydrolases that catalyze the dephosphorylation of threonine and serine remains [14]. While previous studies indicated that the catalytic cycle and mechanism of PPPs depended upon its crystal structure [15,16], Zhang et al [17] reported the structural similarities of the active sites between PPPs and PAPs, suggesting that PAPs might share the catalytic dephosphorylation mechanism with PPPs. Even though sweet potato PAP had shown phosphatase activity toward eight substrates such as ATP, ADP, AMP, NADP, glucose-6-phosphate, sucrose-6-phophate, fructose-6-phosphate, and p-NPP, studies on the dephosphorylation of sweet potato PAP toward relatively large phosphoproteins are limited [18]. The objective of this study was to investigate the dephosphorylation of bacterial flagellin protein by sweet potato PAP, and the effect of sweet potato PAP on the flagellin-mediated inflammatory response in the A549 lung epithelial cell line.
MATERIALS AND METHODS
Reagents and cell culture
Purified flagellin from P. aeruginosa (PA FLA) encoded by the fliC gene [19], and sweet potato PAP were purchased from InvivoGen (Sandiego, CA, USA) and Sigma-Aldrich (St. Louis, MO, USA), respectively. Both were reconstituted in endotoxin-free water (Sigma-Aldrich, USA). PiColor Lock phosphate detection reagent was purchased from Novusbio (Centennial, CO, USA) while the Cymax Human IL-8 ELISA kit was obtained from Ab Frontier (Seoul, Korea). The nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) p65 (Total) InstantOne ELISA kit was procured from Invitrogen, Thermo-Fisher Scientific (Carlsbad, CA, USA).
The human alveolar carcinoma epithelial cell line (A549 cells) was procured from American Type Culture Collection (ATCC CCL-185) and was maintained at F-12K Medium Ham’s F-12K (Kaighn’s) Medium (Gibco Life technologies; Gaithersburg, MD, USA) with 10% fetal bovine serum (Gibco Life technologies, USA) and 1% penicillin-streptomycin (Gibco Life technologies, USA) at 37°C in a 5% CO2 humidified incubator.
HEK-Blue hTLR5 cells (Invitrogen, Thermo-Fisher Scien tific, USA) were cultured at 37°C in Dulbecco modified eagle medium (DMEM; Gibco Life technologies, USA) with 30 μg/mL of blasticidin (Invitrogen, Thermo-Fisher Scientific, USA) and 100 μg/mL of Zeocin (Invitrogen, Thermo-Fisher Scientific, USA).
Dephosphorylation of flagellin by sweet potato PAP at various pHs and temperatures
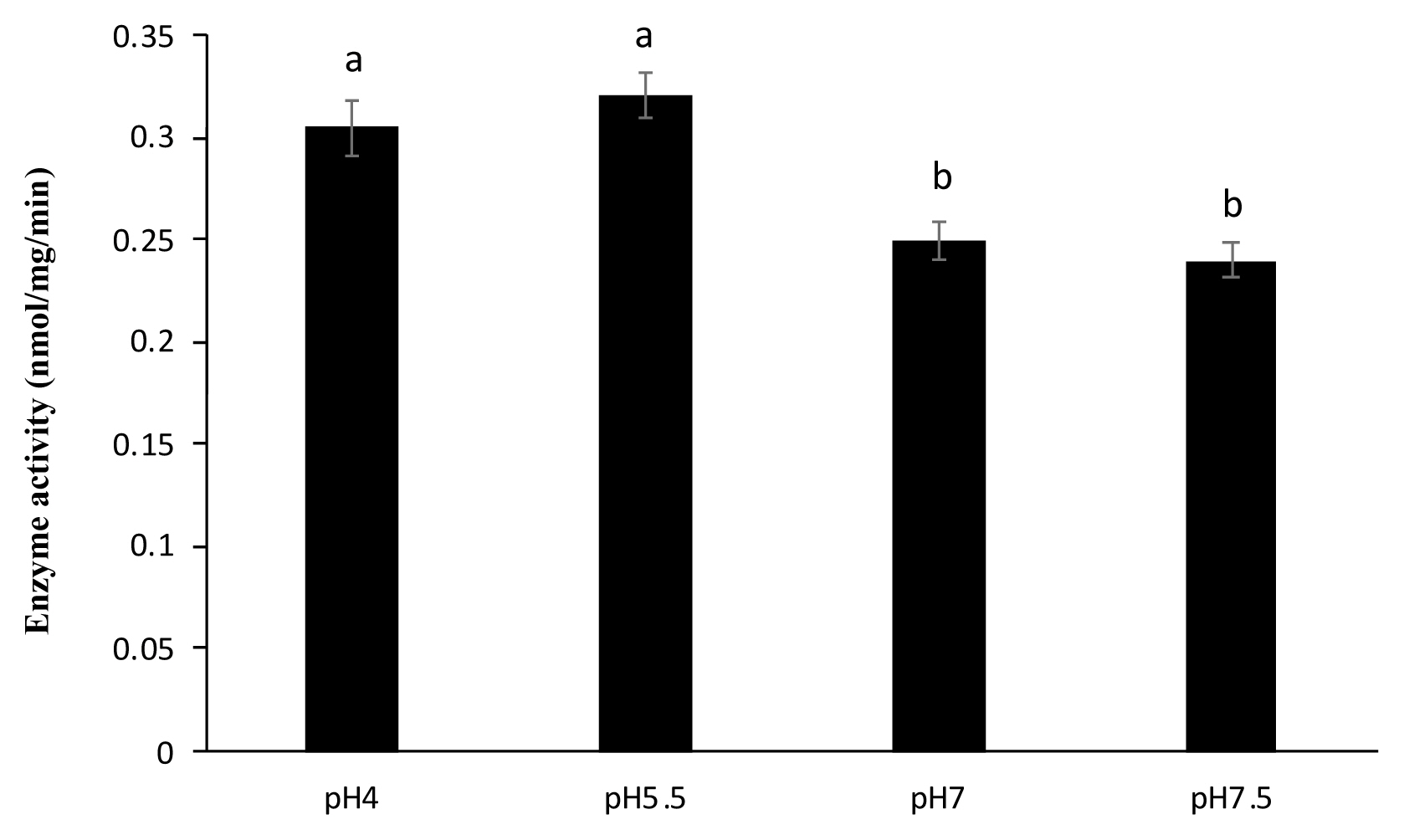
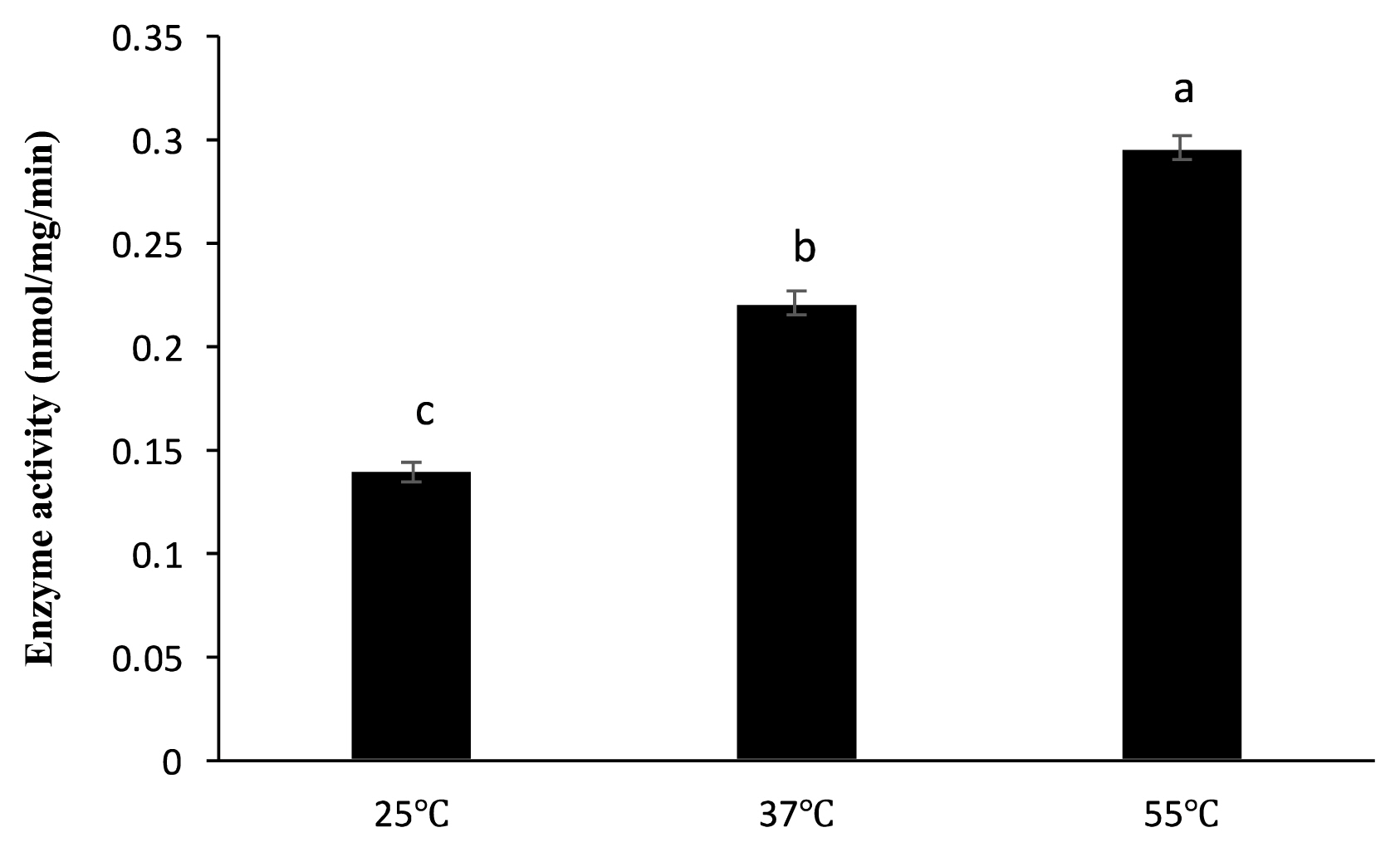
The phosphatase assay was performed at 37°C for 4 hours in a 1-mL reaction mixture containing 4 μL of the enzyme (46.4 μg), 1 μg of PA FLA substrate, and 50 mM Na-acetate (pH 4.0 and 5.5) or Bis-Tris (pH 7.0) or Tris-HCl (pH 7.5). The assay was also performed using the same protocol at conditions of pH 5.5 (Na-acetate) for 4 hours at 25°C, 37°C, and 55°C. The released inorganic phosphates (Pi) were read at optical density (OD) 635 nm, using PiColorLock detection kit (Novusbio; Centennial, CO, USA) following the manufacturer’s instructions. Enzyme activity was defined as the amount of enzyme required to liberate 1 nmol of inorganic phosphate per min under the assay conditions divided by mg of the protein.
Effect of sweet potato PAP on flagellin-induced pro-inflammatory mediators in A549 cells
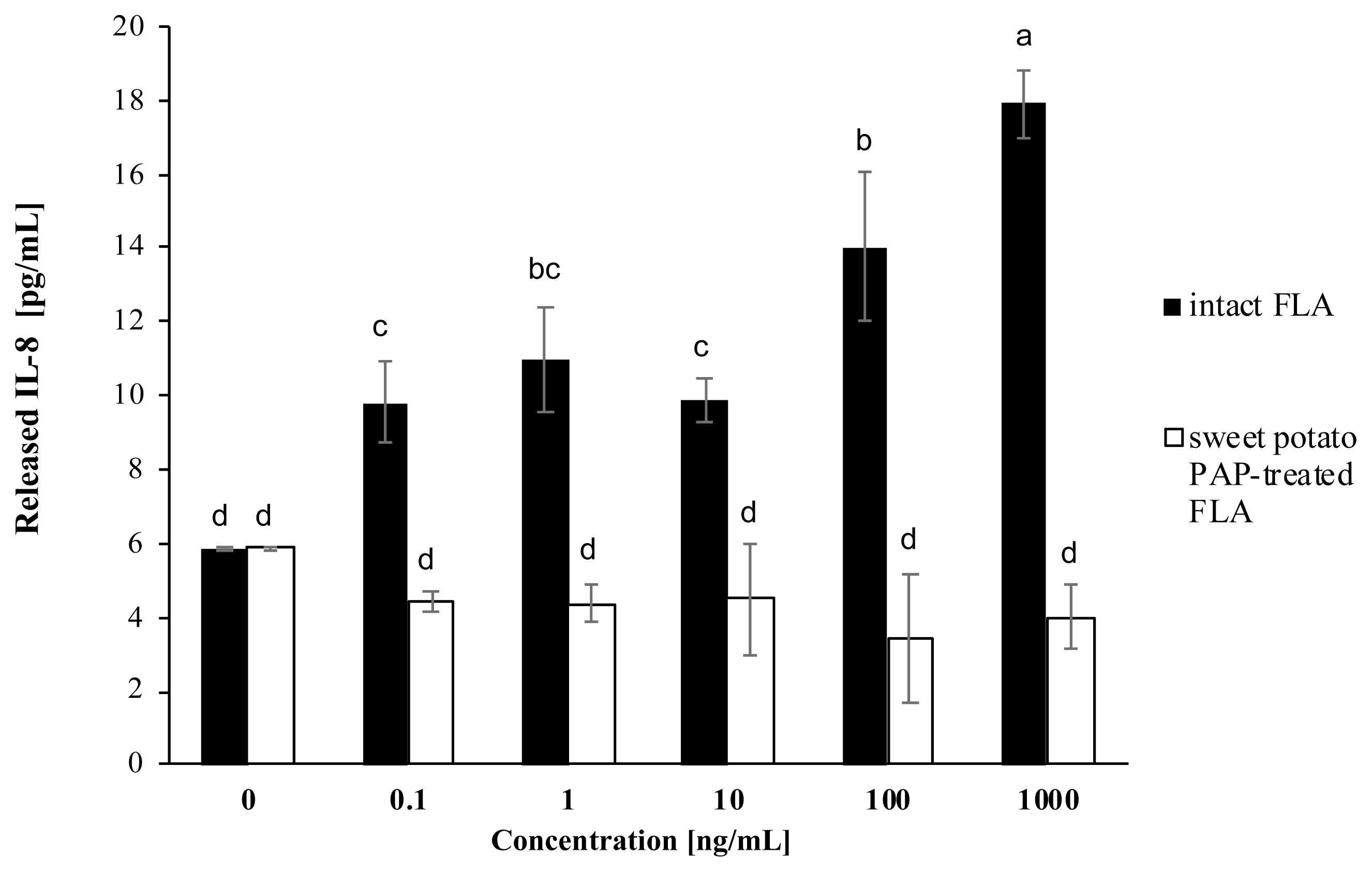
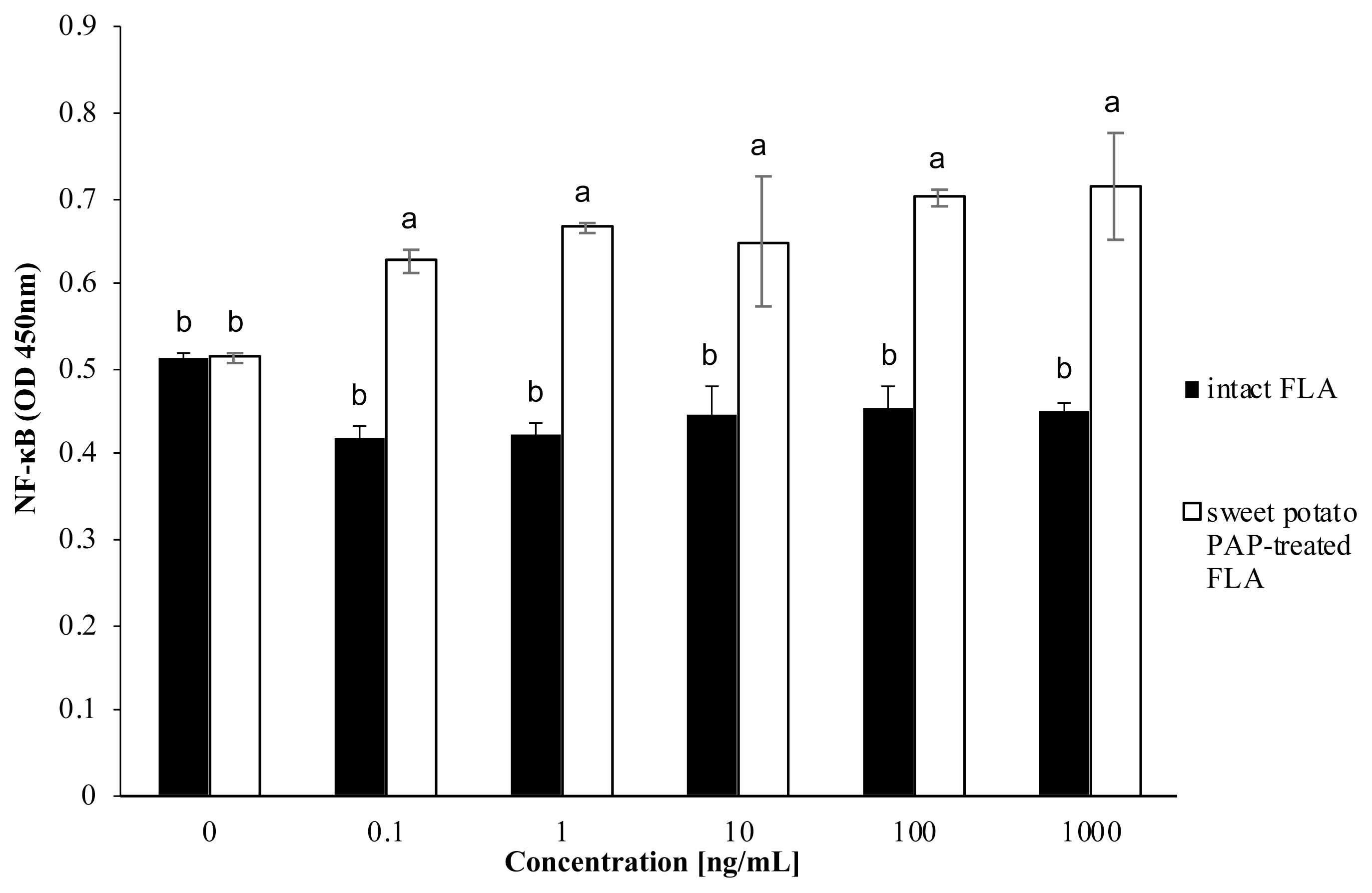
A549 cells were initially seeded onto 96-well plates at a density of 1.5×104 cells per well and cultured to 80% confluency at 37°C in a 5% CO2 incubator. Various concentrations of FLA (0, 0.1, 1, 10, 100, 1,000 ng/mL) were treated with 1 μL of sweet potato PAP (11.6 μg) for 6 h. Then, enzyme-treated FLA and intact FLA at each concentration were applied to the cells for 12 h. The levels of interleukin-8 (IL-8) and NF-κB were assayed at OD 450 nm using Cymax Human IL-8 ELISA kit and NF-κB p65 (Total) InstantOne ELISA Kit, respectively, according to the manufacturer’s instructions.
Effect of sweet potato PAP on flagellin-induced TLR5 activation in TLR5 overexpressing HEK-293 cells
Human TLR5-overexpressing HEK-293 cells (HEK-Blue hTLR5 cells) were cultured in 96-well plates at an initial concentration of 1.5×104 cells per well until 80% confluency. Different concentrations of PA FLA (0, 0.1, 1, 10, 100, 1,000, 5,000 ng/mL) were treated with 1 μL of sweet potato PAP (11.6 μg) for 6 hours. Then, enzyme treated FLA and intact FLA at each concentration were added to the cells. Activated toll-like receptor 5 (TLR5) was monitored at OD 620 nm using the secreted alkaline phosphatase-based assay according to the manufacturer’s protocols.
Statistical analysis
Statistical analyses were conducted using one-way analysis of variance (PROC GLM; SAS 9.4, SAS Inst. Inc., Cary, NC, USA) to search for significant differences between the treatments with Duncan’s multiple range test. The probability level used for statistical significance was p<0.05. The results were presented as the means and standard errors from three experiments.
RESULTS
Effect of sweet potato PAP on PA FLA-induced IL-8 secretion in A549 cells
As shown in Figure 3, PA-FLA stimulated the release of IL-8 in A549 cells. However, PA-FLA treated with sweet potato PAP inhibited the secretion of IL-8 in the cells, decreasing the IL-8 release to about 3.5-fold compared to that of intact PA-FLA, even at 1,000 ng/mL of substrate.
Effect of sweet potato PAP on PA FLA-induced NF-κB activation in A549 cells
Figure 4 showed that PA FLA dephosphorylated by sweet potato PAP repressed the activation of NF-κB in A549 cells compared to intact PA FLA, but there was no significant difference between the substrate levels.
Effect of sweet potato PAP on PA FLA-induced TLR5 activation in TLR5 overexpressing HEK-293 cells
As shown in Figure 5, the activation of TLR5 by PA FLA was effective in TLR5-overexpressing HEK293 cells at substrate concentrations over 100 ng/mL and was the highest at a substrate concentration of 5,000 ng/mL, where PA FLA treated with sweet potato PAP strongly repressed the activation of TLR5.
DISCUSSION
P. aeruginosa infections cause animal diseases and pose a great threat to animal husbandry. The bacteria is responsible for various respiratory diseases, mastitis, otitis, and many others [1,3]. However, the frequently used antibiotics have allowed these bacteria to acquire antibiotic resistance [20,21]. Thus, the importance of finding new alternatives targeting antibiotic resistant bacteria like P. aeruginosa has grown. Although flagellin had been reported to be a PAMP in P. aeruginosa, there are insufficient studies regarding the inactivation of PA FLA.
Several studies on the functions of mammalian PAP have already been conducted and suggested that PAP was involved in physiological events like osteoclastic bone resorption and erythrophagocytosis through the dephosphorylation of certain proteins [22]. Nevertheless, a precise study on the dephosphorylating activity of sweet potato PAP with relatively high molecular weight substances like proteins has not yet been reported. In the present study, sweet potato PAP successfully exhibited phosphatase activity against PA FLA (Figures 1, 2), which was a novel finding. Moreover, sweet potato PAP was highly active at acidic pH range (4 to 5.5) (Figure 1), which was compatible with the result of chickpea PAP (CaPAP7), verifying the typical property of acid phosphatase [23]. Previously, the phosphatase activity of sweet potato PAP for the substrate, p-nitrophenyl phosphate (pNPP) exhibited the highest activity at 50°C [18], which is almost similar to our result of 55°C for PA PLA (Figure 2).
Indeed, bacterial flagellins elicit pro-inflammatory cyto kines such as IL-6 and IL-8 in various cell types [24–27]. As shown in Figure 3, PA FLA provoked the secretion of the universal inflammatory marker IL-8 in A549 cells. However, treatment of the cells with every concentration of substrate dephosphorylated by sweet potato PAP reduced the secretion of IL-8, suggesting that the enzyme exerted anti-inflammatory activity. To some extent, the dephosphorylation of flagellin appears to be related to dysfunction. For example, dephosphorylation of the profission Drp1 (dynamin-related protein 1) protein by the cytosolic phosphatase calcineurin led to the fragmentation of depolarized mitochondria during the cell cycle, differentiation, and death [28].
Moreover, PA FLA dephosphorylated by the enzyme re pressed the activation of NF-κB (Figure 4), which is widely known to mediate IL-8 gene expression [29]. Bacterial flagellin provokes inflammation via stimulating the NF-κB pathway [30,31].
TLRs regulate the innate immune response and contribute to enhancing antibacterial defenses in host cells [32]. Among the TLRs, TLR5 is well-known to recognize FLA on the bacterial cell surface, eliciting inflammatory signaling by activating the NF-κB pathway [31]. When a bacterial infection takes place, host cells over-produce TLR5 to defend against such attacks [32].
As shown in Figure 5, intact PA FLA clearly induced the activation of TLR5 in TLR5- overexpressing HEK-293 cells at higher doses of more than 1,000 ng/mL, which was in good agreement with the previous result observed in 293T cells exposed to purified Salmonella typhimurium flagellin [33], but PA FLA dephosphorylated by sweet potato PAP repressed it. Bacterial flagellins structurally possess two highly conserved N-terminal and C-terminal domains and one central hypervariable domain [32]. In the case of Salmonella enterica-derived flagellin, one variant with a deletion in the N-terminus (FliCΔ1–180) failed to stimulate TLR5 [32]. Presumably, phosphotyrosine exists within the N-terminus of PA FLA and the dephosphorylation of the residue inhibited the interaction of PA FLA with TLR5 [12].
Just as exogenous bovine intestinal alkaline phosphatase has been extensively applied to clinical human trials [34], the conception of the administration of sweet potato PAP by oral delivery or by intravenous injection can be positively suggested to maintain good health for farm and companion animals against the gastrointestinal inflammatory disorder and antibiotics-related infections [34,35]. In conclusion, sweet potato PAP has the potential to be a new alternative agent against the increased antibiotic resistance of P. aeruginosa and may be a new conceptual feed additive to control unwanted inflammatory responses caused by bacterial infections in animal husbandry.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







