





INTRODUCTION
After strong artificial selection for over 150 years, morphological variation has been created in dog populations and more than 400 canine breeds are currently registered worldwide by the Fédération Cynologique Internationale (FCI), and other federations such as the American and British Kennel Clubs [1]. With over 470 million dogs being kept as companion animals worldwide, they were ranked as the leading type of pet in 2018 [2]. In Taiwan, according to survey statistics from the Council of Agriculture, the number of dogs being bred by the end of 2017 had reached 1.78 million. The overall sales revenue of pet-related industries has increased from 15.5 billion New Taiwan dollars (NT$) in the past 10 years to 26.6 billion NT$ (Statistical Bulletin, Ministry of Finance, Republic of China; http://service.mof.gov.tw/File/Attach/86088/File_21588.pdf). The previous roles of companion animals, such as hunting, security, and assistance, have gradually shifted towards them being regarded as family members. With the improvement of the status of pets, owners are increasingly paying attention to their quality of life. Therefore, the demand for various goods and services aimed at companion animals is also increasing. In the early stage of the Taiwan dog-sales market, the characteristics prioritised included the ability to guard, search, attack, and hunt; more recently, these ideals have been superseded by the need to provide companionship and prestige, which have made a recognized pedigree a major factor for buyers. In Taiwan, some legal purebreed grounds used closed flock breeding to keep their dog broodlines, including those three breeds in this study. Especially the Beagles, which have been used to hunt hares in the British Isles for centuries, and which were brought to the United States in 1880 to breed in large numbers. The modern Beagles have been modified to become a pet dog, and were often used as experimental dogs [3]. It has considerable medical research value, and this dog was also cultivated as laboratory animal for medical research in Taiwan.
So far, many dog breeds have been developed in order to meet appearance standards and maintain the purity of their bloodlines. Breeding companies usually adopt inbreeding methods, which can lead to the occurrence of many genetic diseases. Generally, in the natural state—unlike with experimental animals for which it can be essential to reduce individual differences for study purposes—inbreeding approaches have seldom been used, in order to avoid inbreeding depression [4]. The Royal Society for the Prevention of Cruelty to Animals (RSPCA) pointed out that dogs are now subject to more than 300 genetic diseases. Not only do the animals have to bear great suffering, but their owners also experience mental pressures and financial losses [5]. Therefore, in order to maintain high heterozygosity and stability of the genetic background of the entire population, it is necessary to have a reliable breeding system and genetic monitoring [6]. However, in Taiwan, the genealogical and registration data requirements for many breeds of dog are incomplete or remain to be established, and most of the certificates of pedigree produced by breeding sites in the market lack the backing of publicly trusted authorities, so trading disputes arise from time to time. Although trading in companion animals is discouraged in many countries today, the market in Taiwan is booming. In order to prevent companion animals from being afflicted by genetic diseases, in addition to promoting care by and education of owners, an important factor is reducing inbreeding. The Kennel Club has established a breeding certification system for 68 breeds since 2008, with the total number now reaching 222 breeds. Breeders can inquire about the diseases to be screened for each dog and the procedures for obtaining certification. After the puppies are certified by the Kennel Club, the breeders are issued with a puppy sale wallet; this measure not only protects the profits of the owner, but also reduces disputes over the sale of companion animals [7].
Regarding the research on dog microsatellite markers, a lot of information has been discovered in conjunction with the elucidation of DNA sequences [8]. As early as the 1990s, there have been many studies on dog microsatellite markers [9,10]. The application of canine microsatellite markers in modern times has mainly focused on two fields: one is evaluating the genetic structure polymorphism of populations, and the other is proving a platform for individual identification or paternity [11,12]. Wictum et al [13] searched for published dog genome sequences, and selected suitable microsatellite markers based on the stability and high polymorphism that were required for forensic applications. A total of 15 microsatellite markers and a marker related to gender comprise the multiplex system, DogFiler [13]. This is the first dog-identification data system created based on the recommendations of the Scientific Working Group for DNA Analysis Methods (SWGDAM) in the United States. At present, DogFiler has been integrated into forensic casework, and is widely accepted by courts in the United States. Owing to the relatively long period of strong artificial selection, the differentiation between dog breeds has been large. The sequences on both sides of microsatellite markers may have different degrees of variation, making it impossible to perform polymerase chain reaction (PCR) amplification in similar breeds. Even if the amplification is successful, the number of alleles and polymorphisms may be far lower than in the original breeds [14]. Therefore, when conducting research on the population genetics of specific breeds of dog, it is necessary to develop new microsatellite markers.
To date, only a few domestic biotechnology companies in Taiwan use foreign dog microsatellite commercial kits, such as the StockMarks for dogs genotyping kit, for individual genetic analysis or genetic structure analysis at dog breeding sites. So, are they applicable to Taiwan? To our knowledge, there are no relevant published reports on existing dog breeds for reference and analysis. Therefore, the development of microsatellite markers suitable for the companion animal population in Taiwan is a crucial task to establish a molecular-detection platform for domestic dogs. Moreover, such a platform will make it possible to evaluate the inbreeding level of the dog population in Taiwan. In addition to assisting with the formulation of domestic dog breeding-management policies, this could also enhance Taiwan’s positive governance perspectives on animal protection and animal welfare.
MATERIALS AND METHODS
Experimental animals and sample collection
In total, five populations of dogs from legal breeding ground were investigated in this study. Blood samples were collected from 113 individuals belonging to these three populations which were popular in Taiwan: 17 bichons (BIs); 14 schnauzers (SCs); 74 Taiwanese beagles (BETs); and eight Japanese beagles (BEJs), two of which were distinguished as group A (BEJAs) and six of which were distinguished as group B (BEJBs) (Table 1). For each dog, 5 mL of blood was drawn from the jugular or cephalic vein. Whole genomic DNA (gDNA) was then extracted with the Genomic DNA Isolation Reagent (GenePure Technology Co., Ltd, Taichung, Taiwan) using the standard phenol-chloroform method. NanoDrop 2000c (Thermo Fisher Scientific Inc., Waltham, MA, USA) detected the DNA concentration and the optical density (OD) 260/280 values between 1.8 and 2.0, and samples were stored in a refrigerator at −20°C for later use. All animal experiments were approved by the Institutional Animal Care and Use Committee of our university (Protocol number: NTU Animal Experiments No. 2012-089).
Isolation of microsatellite loci
The gDNA was isolated from blood samples taken from one male and one female BI using the following steps, which were slightly modified from the procedure described by Glenn and Schable [15]. The isolated gDNA was partially digested with RsaI and XmnI (NEB, Ipswich, MA, USA) until most of the DNA fragments were between 300 and 1,000 base pairs (bp) in length. Re-naturing of single-strand SuperSNX24 forward (5′GTTTAAGGCCTAGCTAGCAGAATC3′) and SuperSNX24+4P reverse (5′pGATTCTGCTAGCTAGGC CTTAAACAAAA3′) formed SuperSNX24 linkers that were ligated to the digested DNA fragments. Linked DNA fragments were then amplified with SuperSNX24. Next, microsatellite markers including fragments with biotinated probes were isolated and enriched. The following three probe compositions were used: i) (TG)12, (ACT)12, (ACTG)6, and (ACAG)6; ii) (AG)12, (ACAT)8, (AACT)8, and (AAGT)8; and iii) (AAG)8, (AAAC)6, (AATC)6, and (AGAT)6. The biotinated probes were then annealed to fragments of gDNA containing complementary regions. Finally, the microsatellite-containing fragments were enriched using streptavidin-labeled metal beads (Dynabeads M-280 Streptavidin, catalog #11205D; Invitrogen, Waltham, MA, USA). The enriched segments were then cloned into pGEM-T Easy vector (pGEM-T Easy Vector system; Promega, Madison, WI, USA) and sequenced with an ABI 3730XL DNA Analyzer (Applied Biosystem PRISM, Waltham, MA, USA). The fragments with higher numbers of repeats were selected for polymorphism testing.
Polymerase chain reaction and polymorphism testing
The selected highly repeated fragments were subjected to PCR and polymorphism testing, which would verify whether the microsatellite loci could be amplified and used to show diversity in the investigated populations. Primers for loci amplification were designed using Primer3plus. CAG-tag (5′-CAGTCGGGCGTCATCA-3′) or M13Reverse (5′-GGAA ACAGCTATGACCAT-3′) was added to the 5′ end of one of each primer pair. Following the protocol described by Schuelke [16], a fluorescent dye-labeled tag, as a third primer, was used with the primer pair to amplify the target fragments that were detectable upon capillary electrophoresis. Eight dog samples from four BIs and four SCs were tested at a 20-μL volume using a thermalcycler (GeneAmp PCR system 9700; Applied Biosystems, USA) containing 0.5 U Taq DNA polymerase (TAKARA, Kusatsu, Shiga, Japan), 1×PCR buffer (1.5 mM MgCl2), 0.2 mM deoxyribose nucleotide triphosphate (dNTP), 0.2 μM unlabeled primer, 0.04 μM tag-labeled primer, 0.16 μM dye-labeled tag, and 50 ng gDNA. The PCR cycling program was as follows: 95°C for 5 min, 35 cycles of 95°C for 30 s, 50°C to 65°C for 40 s, 72°C for 40 s, and a final elongation at 72°C for 7 min. The amplified microsatellite PCR products were analyzed with a DNA analyzer (ABI PRISM 3730 DNA analyzer; Applied Biosystems, USA). Allelic sizes of all loci were estimated relative to the in-line GeneScan500 LIZ Size Standard marker (ABI PRISM; Applied Biosystems, USA). The fragment size was calibrated and analyzed with Peak Scanner Software version 1.0 (ABI PRISM; Applied Biosystems, USA). Those loci that had an allele number greater than two and similar annealing temperatures were selected for whole-population analysis.
Multiplex polymerase chain reaction
In total, 14 microsatellite loci were selected (Table 2). The 5′ end of each forward primer was labeled with the fluorescent dyes fluorescein (FAM), 2′-chloro-7′phenyl-1,4-dichloro-6-carboxy-fluorescein (VIC), red color fluorescent (PET), or 2′-chloro-5′-fluoro-7′,8′-benzo-1,4-dichloro-6-carboxyfluorescein (NED). Multiplex PCR was performed on a 30-μL reaction containing 1 U Taq DNA polymerase (TAKARA, Japan), 1×PCR buffer (1.5 mM MgCl2), 0.3 mM dNTP, 0.3 μM forward and reverse primer, and a 100-ng DNA template. The steps of the PCR program and genotype detection were as described above.
Statistical analysis
For each locus and population, and across populations, commonly derived statistics from the microsatellite genotypic data, including allele frequencies, the observed number of alleles (No), the observed heterozygosity (Ho), the expected heterozygosity (HE), and the polymorphic information content (PIC), were calculated with the Microsatellite Toolkit. The Hardy–Weinberg equilibrium test was performed using the GENEPOP computer program [17], which also was used to estimate F-statistics (the global mean inbreeding coefficient [FIT], the average inbreeding coefficient of an individual with respect to the local subpopulation [FIS], and the average inbreeding coefficient of subpopulations relative to the total population [FST]) for each locus, the pairwise FST between populations, and the average inbreeding coefficient (FIS). Nei’s genetic distance (DA) [18] between populations was measured with the Microsatellite Analyzer [19]. A phylogenetic tree was generated via the PHYLIP program [20] using the neighbor-joining (NJ) method with a bootstrap test of 1,000 resampling of loci with replacement [21].
The model-based approach proposed for the population structure analysis of the dog populations was carried out with the software STRUCTURE 2.3.1 [22], which was used to assess the genomic clustering (K) of the sample. To obtain a representative value of K for data modeling, 10 independent runs were performed for each value from one to seven. The run length was set to 100,000 burn-ins followed by 100,000 iterations. In addition, a principal coordinate analysis (PCoA) three-dimensional (3D) map drawn by GenAlEx [23] was used to confirm the inter-population situation.
The effectiveness of individual identification is expressed by the probability of identity (P(ID)), which is the probability that two individuals are randomly selected from the population and the genotypes of the two are identical at a single locus. The theoretical expectations are as follows:
where pi and pj are the frequencies of the ith and jth alleles, and i ≠ j. When the molecular markers are co-dominant, such as microsatellite markers, and the identified individuals have close kinship, the following probability of identity among sibs (P(ID)sib) is applied:
RESULTS
Polymorphism, heterozygosity, and F-Statistics of novel microsatellite loci
The 14 microsatellite markers were used to perform genotyping on 113 samples from the three dog breeds (SCs, BIs, and BETs/BEJs). Polymorphism was clearly observed at most of the microsatellite loci in the three breeds. The genetic characteristics of the 14 microsatellite loci are listed in Table 3. The average number of alleles per locus (Na) was 6.3. The actual number of alleles ranged from 2 (SEL093 and SEL094) to 13 (SEL115). The average number of effective alleles per locus (Ne) ranged from 1.4 (SEL094) to 7.6 (SEL005), with an average across loci value of 3.6. The PIC value ranged from 0.249 (SEL094) to 0.855 (SEL005), with an overall average value of 0.612. All of the selected microsatellite loci in this study were sufficiently polymorphic, indicating that they were suitable for the genetic analysis of dogs.
The HE among the 14 microsatellite loci ranged from 0.293 (SEL094) to 0.873 (SEL005), with an average of 0.662. The HO among the 14 microsatellite loci ranged from 0.248 (SEL094) to 0.814 (SEL005), with an average of 0.567 (Table 3). However, there were eight loci—namely, SEL025, SEL030, SEL031, SEL035, SEL068, SEL098, SEL115, and SEL118—that significantly departed from the Hardy–Weinberg equilibrium (p<0.01).
The Wright’s F-statistic values (FIS, FIT, and FST) for each locus are shown in Table 3. The average FIS for all the loci was 0.002, and the FIS per locus varied from −0.191 (SEL093) to 0.328 (SEL115). The average FIT for all the loci was 0.209, and the FIT per locus varied from −0.070 (SEL034) to 0.506 (SEL115). The mean FST for all the loci was 0.212. This value implied that around 21.2% of the total genetic variation was caused by population differences and that 78.8% of the total genetic variation was due to genetic differentiation among the individuals within each population.
Intra–population genetic variability
The genetic statistics relating to polymorphism, including HE, HO, PIC, the mean observed number of alleles, and the mean effective number of alleles, were calculated to estimate the allelic diversity at each locus of the population. These genetic parameters across the 14 loci for the three dog populations are listed in Table 4. The HE varied from 0.480 (SCs) to 0.624 (BEs), whereas the HO varied from 0.485 (SCs) to 0.587 (BEs), and the PIC ranged from 0.407 (SCs) to 0.567 (BEs). The SC population had the lowest values of HO, HE, and PIC.
Among the three breeds, the BE population had the highest observed mean number of alleles (MNA) (5.3), followed by the BI (4.0) and SC (3.3) populations, while the latter had the smallest observed MNA. Negative FIS values were observed in the BI and SC populations, indicating an insufficient degree of inbreeding. The deviation from the Hardy–Weinberg proportions within populations (FIS) varied from −0.032 to 0.045. The highest inbreeding effects were found in the BE population (0.045), which significantly deviated from the Hardy–Weinberg equilibrium (p<0.01) (Table 4) in comparison to the other two populations.
Inter-population genetic variation
To estimate genetic variation among the five dog populations, two parameters—FST and genetic distance (D) calculated by the gene frequency of each animal at each microsatellite locus—were evaluated in this study. The values of FST and D for each test population pair are summarized in Table 5. The FST for each population pair was highly significant (p< 0.05). The FST values of the population pairs varied from 0.165 (for the BET and BEJB population pair) to 0.405 (for the SC and BEJB population pair). The genetic distances between the dog population pairs varied from 0.400 (for the BET and BEJB population pair) to 0.993 (for the SC and BEJA population pair). Surprisingly, the second relatively low genetic distance and FST value were not between other BE populations. The genetic distances and FST values of the BI–BET and SC–BET population pairs were less than the BEJA–BEJB and BEJA–BET population pairs.
Population differentiation analysis
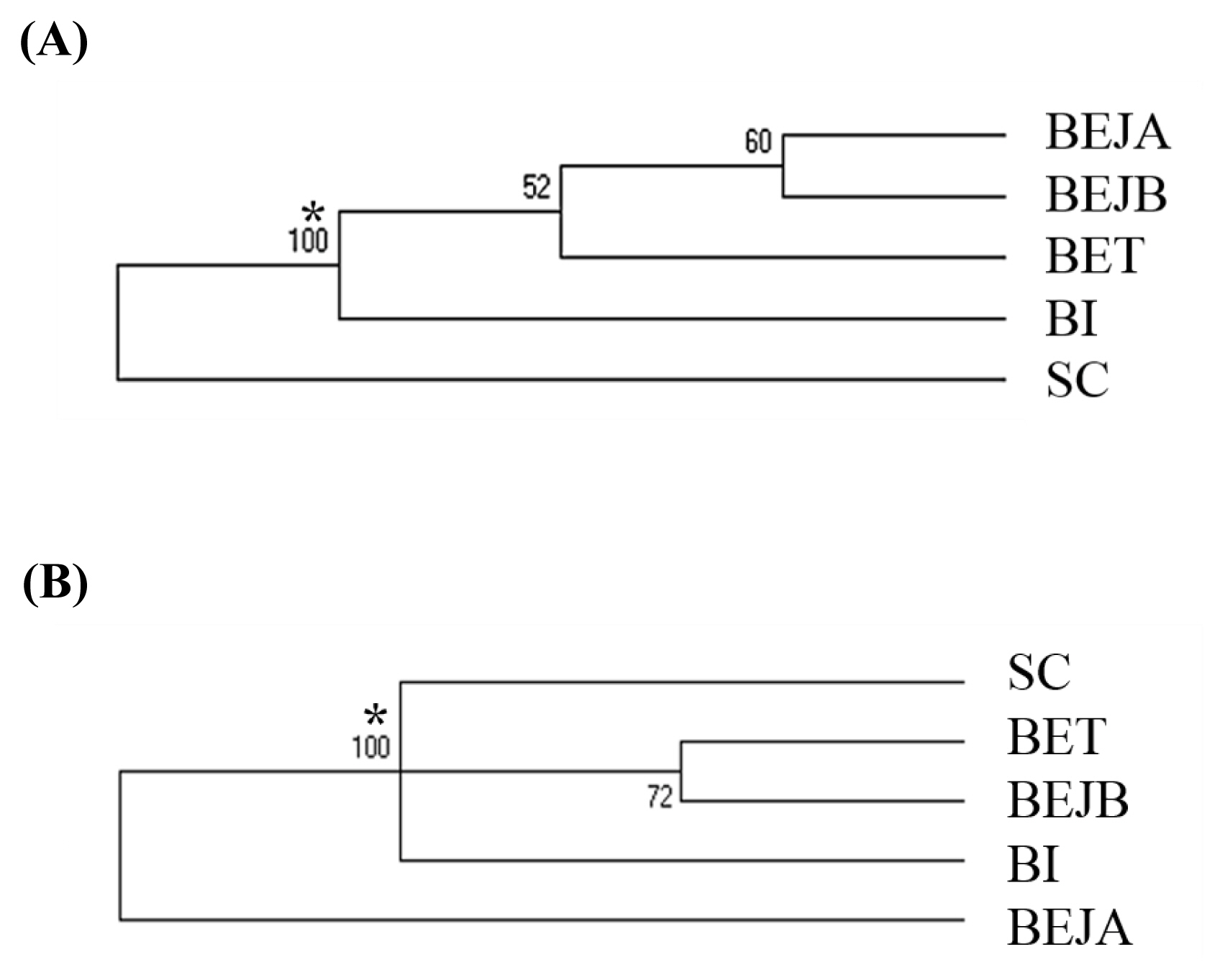
The Nei’s standard genetic distance of these three populations of dogs in Taiwan was calculated. A DA distance matrix was used to build an individual phylogenetic tree with the NJ method (Figure 1). The results showed that the individual phylogenetic tree could be divided into three main clusters: SC, BI, and BE. Among these, the BE cluster could be further divided into a Taiwan population (BET) and two Japanese sub-populations: a Japan A (BEJA) and Japan B (BEJB) population. A phylogenetic tree using the NJ method with bootstrap resampling (n = 1,000) of the 14 microsatellite loci was constructed with the PHYLIP software. In the NJ tree (Figure 2A), the entire dog population could also be divided into three main clusters: SC, BI, and BE. The results of the main clusters were consistent with the results of the individual phylogenetic tree (Figure 1). In addition, the bootstrap value between the SC and other dog populations was 100%, which showed that the genetic distance between these groups was relatively large. Unlike the NJ method, the phylogenetic tree constructed by the unweighted arithmetic average pair group (UPGMA) method (Figure 2B) showed that the BEJA group was clearly distanced from others, with a bootstrap value of 100%. The remaining dog groups were divided into three clusters: SC, BI, and BET and BEJB.
A PCoA of pair-wise genetic distances among the five examined dog sub-populations was used to represent the relative positions of the populations. The first (PC1), second (PC2), and third (PC3) PCos accounted for 44.15%, 26.35%, and 19.97% of the total variation, respectively (Figure 3). The distance between the BEJA and BEJB groups was relatively close. The distance between the BET group and both the BEJA and BEJB groups was closer than the distance between the BET group and both the SC and BI groups.
Population structure analysis
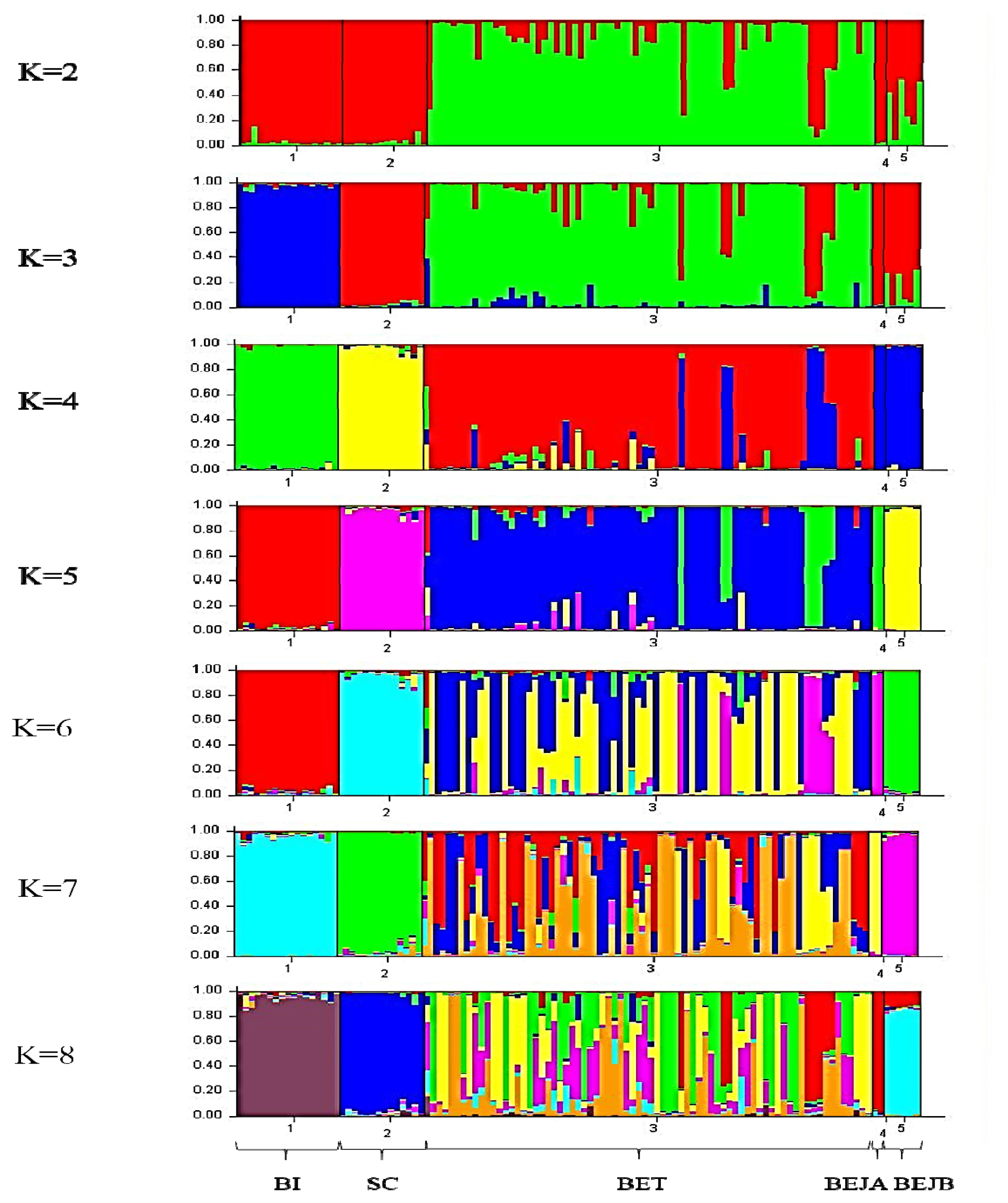
The STRUCTURE software program using Bayesian model-based clustering algorithms of multi-locus genotypes was utilized to assign individuals to populations via estimated individual admixture proportions and to infer the number of populations (K) for a given sample. The results of the analysis are shown in Figure 4. It was mainly divided into Taiwan Beagle population and other populations at K = 2. When K = 3, the BI group was separated. At K = 4, it is mainly divided into BI, SC, BET, and BEJ four clusters. Until K = 5, the two BRJ clusters are separated finally. When the K value over 5, the BET populations are further subdivided into different clusters.
Probability of identity and power of exclusion
The analysis of probability of identity involved three dog populations, respectively, and all populations together was calculated by the probability of identity (P(ID)) and the identification rate of close relatives (P(ID)sib) of the 14 single new microsatellite markers. The results are listed in Table 6 and Table 7. The P(ID) of BE populations per locus varied from 0.042 (SEL005) to 0.491 (SEL094). The combined P(ID) values for all loci was 3.7×10−11. The P(ID) of each locus in BIs ranged from 0.060 (SEL005) to 1.000 (SEL094). The combined P(ID) values for all loci was 7.8×10−9. The P(ID) of SC populations per locus varied from 0.097 (SEL005) to 0.869 (SEL115). The combined P(ID) values for all loci was 1.2×10−7. In addition, the P(ID) of the 14 novel microsatellite loci for all dog populations ranged from 0.031 (SEL005) to 0.843 (SEL094), and the comprehensive P(ID) was 1.7×10−12.
When individuals in the population to be explored have close relatives, such as full-sib or half-sib, it is more appropriate to evaluate the proportion of identification using the P(ID)sib. The P(ID)sib of BE populations per locus varied from 0.337 (SEL005) to 0.701 (SEL094). The combined P(ID)sib values for all loci was 3.9×10−5. In BIs, the P(ID)sib ranged from 0.358 (SEL005) to 1.000 (SEL094), and the combined P(ID)sib values for all loci was 2.6×10−4. The P(ID)sib of SC populations per locus varied from 0.403 (SEL005) to 0.933 (SEL115). The combined P(ID)sib values of SC for all loci was 7.1×10−4.
In the paternity test for dogs, according to the formula of power of exclusion (PE) proposed by Jamieson [25], the value calculated using the 14 new microsatellite markers’ allele frequencies was 99.98%. This meant that when the genotypes of the mother and offspring were known, the possibility of being a biological father could be almost completely eliminated for an individual who was not a sire.
DISCUSSION
In this study, 14 sets of novel microsatellite markers were developed and used to analyze the genetic variation of three dog breeds—SC, BI, and BE. The results showed that the Na of the 14 novel microsatellite loci was 6.3, and the Ne was 3.6 (Table 3). The Na of locus SEL115 was 13, whereas the Ne was only 3.8. The reason might have been that the distribution of allele frequency was mainly concentrated on three alleles—243 bp (13.3%), 247 bp (10.6%), and 251 bp (47.4%) (Supplementary Table S1)—such that in some cases the uneven distribution of allele frequencies caused a large gap between the Na and the Ne.
The HE, HO, and PIC are commonly used to assess the polymorphism of microsatellite loci in the analyzed population. The HO refers to the observed heterozygosity of each locus, which represents the actual proportion of heterozygous individuals in the population. The HE is the expected heterozygosity of each locus, which is the expected proportion of heterozygous individuals in the population that is calculated according to the Hardy–Weinberg Law. The PIC is the degree of polymorphism of each locus. Using the 14 novel microsatellite markers to analyze our dog populations, the average HE was 0.662, which showed that the values of most of the microsatellite markers were within high expected heterozygosity (HE>0.5). The average value of HO was 0.567, which also fell within the range of high observed heterogeneity (0.7>HO>0.5) [26]. The average value of PIC was 0.612, which fell within the range of high polymorphic information content (PIC>0.5) [27] (Table 3). The results of this experiment were similar to the study of Radko et al [28] using 18 sets of microsatellite markers to analyze the Polish Tatra shepherd dog. Their results showed that the average HE was 0.643, the average HO was 0.645, the average PIC was 0.598, and the values of the three variables were all greater than 0.5. Therefore, their study indicated that the tested Tatra shepherd dog population was highly polymorphic. In another study [29], eight breeds of dog were surveyed by 21 microsatellite markers. PIC values over 0.5 were measured for 15 markers. The average value of the PIC was 0.555. Compared with these reports, the three variables in the current experiment were highly polymorphic, indicating that the 14 novel microsatellite loci should be able to effectively analyze the genetic structure and genetic variation of the three breeds of dogs analyzed in this experiment.
The HE (0.412 and 0.293) and HO (0.451 and 0.248) values of loci SEL093 and SEL094 were both less than 0.5, as were the respective PIC (0.326 and 0.249) values. The cause of this result, for which there were only two alleles in two loci, was supposed to be sampling error. However, some reports have suggested that the number of alleles for microsatellite markers should be three or more to reduce the standard deviation of distance calculation [30]. The reason why the loci SEL093 and SEL094 were selected in this experiment was that the number of dog breeds analyzed was relatively small. If the number of breeds is increased, perhaps the allele number of these two microsatellite loci could be increased, and the three variables will be likely to increase as well. On this basis, the two microsatellite loci SEL093 and SEL094 were retained as potential canine microsatellite loci in this experiment.
In terms of the analysis of population genetic structure, we applied the FIS, FST, and FIT statistics to evaluate the distribution of genetic variation within and between populations. The average FIS value of the 14 new microsatellite markers was 0.002. This value was positive and low. The percentage of heterozygotes in the overall tested dog population was less than expected—that is, there was an inbreeding phenomenon—but the average value of FIS was around 0.002, which indicated that the situation in the dog population was not serious. The mean FST for all the loci was 0.212. This fell within the range of high differentiation (0.15<FST<0.25), according to the Sewall Wright rules [31], indicating that there was high differentiation among the three breeds of dogs in this study.
Kang et al [32] investigated the genetic structure of local dogs in South Korea and establish an individual and paternity identification system through evaluating the polymorphisms of the populations with three variables: HO, HE, and PIC. Between nine and 11 microsatellite loci were used for genetic analysis of two local breeds from South Korea and three exotic dog breeds in their study. The sample selection criterion was at least one generation of unrelated dog individuals. The results showed that the average HO for each breed ranged from 0.65 to 0.78; the HE ranged from 0.71 to 0.85; and the PIC ranged from 0.66 to 0.82. In this study, the average value of HE for the three varieties ranged from 0.480 (SCs) to 0.624 (BEs), and average HO ranged from 0.485 (SCs) to 0.587 (BEs). The average PICs ranged between 0.407 (SCs) and 0.567 (BEs) (Table 4). Compared with the abovementioned studies, the three variables in our experiment showed slightly lower values, which may have been caused by differences in the individual dogs included in this experiment: some of the animals were blood-relatives and were full-sibs or half-sibs, so their genetic backgrounds were similar, leading to slightly lower polymorphisms. Future testing of individual Taiwanese dogs with different origins or more distant blood relationships could improve the applicability of these new microsatellite markers.
Among the different varieties of FIS, only the average value for BEs (0.045) was positive. The results showed that although BEs in this experiment had high genetic variation, the positive value of FIS indicated that the proportion of heterozygous individuals was still too small to achieve the Hardy–Weinberg balance; Iindeed, it deviated significantly from the expected value (p<0.01). This result may reflect the fact that the fathers of the BE population in this experiment comprised a small number of male dogs, which was not reflective of the situation of mating by chance. The BIs and SCs did not deviate from the Hardy–Weinberg equilibrium, and the FIS values of these two breeds were negative. It can thus be inferred that these two breeds had no inbreeding issues, the genetic backgrounds of the parents were different, and the number of male and female animals was equal [33]. Therefore, it was supposed that the deviation of the entire dog population from the Hardy–Weinberg balance was attributable to the deviation of the BE population.
The individual phylogenetic tree of the dogs (Figure 1) was constructed by the NJ method, and the cluster analysis diagram drawn by the STRUCTURE software (Figure 4), and the dogs in this experiment were divided into five groups: SC, BI, BET, BEJA, and BEJB. In the phylogenetic tree of dog populations drawn by the NJ method (Figure 2A), the distinctive main clusters were consistent with the result of the individual phylogenetic tree: both could clearly distinguish the BI, SC, and BE groups, and the BET and BEJ groups were closely identified. The different breeds of dogs could be clearly differentiated by the microsatellite markers used in this experiment. The differentiation of dog breed in tree would be caused by the unique alleles. For example, the alleles 185 bp and 201 bp of the SEL035 locus were only found in the BI and SC populations, respectively (Supplementary Table S1). In the phylogenetic tree drawn by the NJ method, the bootstrap value of the BET and the BEJ group was 52%, which means that only 52% of the analysis results separated them. When the bootstrap value between the two populations was not greater than 70% in neighbor joining tree, it showed that the clustering of that two populations were not obvious [34]. In the NJ phylogenetic tree, the bootstrap value between the SC and other dog populations was 100%, and the result was in the highly reliable range (bootstrap value>70%). That showed that the genetic distance between the SC and other dog populations was relatively long. However, the UPGMA phylogenetic tree (Figure 2B) shows that the BEJA group was far away from other dog groups, and the bootstrap value was 100%. But the UPGMA method based on constant-rate assumption, and the distance between samples on the same branch is the same, so this method is only suitable for the case where all samples have the same evolution distance [35], so recently it was less used to construct a phylogenetic tree, and the NJ method was a more commonly used method for drawing a phylogenetic tree.
The PCoA draws a 3D stereogram based on the genetic distance between the populations (Figure 3), and distinguishes the genetic distance among the populations by the variance of the three principal coordinate axes. It can be found that the two BEJ groups were relatively close, which was consistent with the close geographical relationship between the two, and the relative distance between the two groups and the BET group was closer than the relative distance between the two groups and the SC and the BI. The results of the NJ phylogenetic tree can also be confirmed in the 3D map of the PCoA. The SC and the BEJA were located on the farthest sides of the 3D map, so there was a maximum genetic distance between the two populations.
In the individual analysis, the combined probability of identification (P(ID)) of the 14 new microsatellite loci for the entire dog population was 1.7×10−12 (Table 6), and according to the survey of the Council of Agriculture up until the end of 2019, the total number of dogs in Taiwan was about 1.54×106 [36]. That meant when the current number of Taiwanese dog populations is analyzed by the 14 new microsatellite markers used in this experiment, the probability of appearing exactly the same genotype is very low. Although the dog population in this experiment contained only three breeds, however, when examining P(ID) of a single dog breed, the credibility is still high. There are no statistics on the numbers of dogs of different breeds currently in Taiwan; however, it can be ascertained that the number of dogs in any single breed cannot exceed the total number of dogs. Therefore, the P(ID) of the three dog populations respectively in this study should cover the total number of dogs in Taiwan in 2019. It was confirmed that the probability of the same genotype being identified in two individuals with 14 the sets of novel microsatellite markers was very low.
The combined probability of identity among sibs (P(ID)sib) was 1.6×10−5 (Table 7) in this study. According to Waits et al [24], their markers were sufficient to identify close relatives of the natural population when the P(ID)sib was between 10−3 and 10−4. The P(ID)sib values of the three breeds of dogs in this experiment generally met this recommended standard. This confirmed that the 14 novel microsatellite markers were applicable in the individual identification of close relatives of BEs, BIs, and SCs in Taiwan.
In the paternity tests of dogs, the PE of the 14 new microsatellite markers in this experiment was 99.98%, which meant that when the genotypes of the mother and offspring were known, individuals that were not the biological father could be ruled out. According to the pedigree data for the BE population collected in this experiment, there was one of the cases where the mother–child genotypes were known, the individual registered as the biological father in the pedigree was compared with the genotypes of 14 novel microsatellite markers and which was not found to provid any of the alleles to the offspring at six of the loci. Therefore, it could be inferred that the male was not the biological father, and suggested that the registration of the dog’s pedigree data was inaccurate. This supports the suggestion that many dog-breeding facilities in Taiwan are still not rigorous enough for pedigree registration. Therefore, using paternity facilities with high PE to improve the registration of pedigree in Taiwan is important. The microsatellite markers developed in this study may be suitable for this purpose to avoid the potential damage caused by inbreeding.
At present study, many countries used microsatellite markers to analyze dog populations. For developing a platform of paternity and individual identification, the American Kennel Club (AKC) analyzed 108 dog breeds using 17 microsatellite markers. The results show that the average HO was 0.60, and the average value of PIC was 0.56. PE was more than 99% in all breeds, and the combined P(ID) was 3.2×10−8. The American Kennel Association considered that 17 microsatellite markers were sufficient for ordinarily paternity identification [37]. Eichmann et al. established Austrian dog DNA profiling for investigation of dog-related accidents and crimes, using 15 sets of highly polymorphic microsatellite markers to analyze 45 dog breeds [38]. The results revealed that the average HO was 0.74, the average PIC was 0.82, and the combined P(ID)sib was 8.5×10−8, showing that these 15 microsatellite markers are sufficient for individual identification of dogs in Austria. Kang et al [32] used 9 to 11 sets of microsatellite markers to detect the genetic structure of local dogs in South Korea and established an individual and paternity identification system to perform genetic analysis of two local breeds of South Korea and three foreign dog breeds. The results showed that the average HO for each breed was between 0.65 and 0.78; the average PIC was between 0.66 and 0.82; and the average PE was more than 99% in all breeds. The average HO value (0.57) of the 14 new microsatellite markers in this study near the range of the research results of the previous countries (0.60 to 0.74). The average value of PIC (0.61) was also within the range of the results of the aforementioned countries (0.56 to 0.82). According to the researches before, the 14 new microsatellite markers developed in this study were highly polymorphic and suitable to analyze the three breeds of dogs in Taiwan.
In conclusion, using 14 novel microsatellite markers to analyze the beagle, bichon, and schnauzer populations in Taiwan, the results showed that their average expected heterozygosity, observation heterozygosity, and polymorphism information content were all at high levels. Therefore, these new microsatellite markers have high applicability to the analyzed populations. These results indicate that the new microsatellite markers have good resolution when applied to the detection of differences among dog breeds. It was confirmed that the opportunity of identifying the exact same genotype among the analysis of the 14 new microsatellite markers was very low. In addition, the power exclusion was enough high to be a good tool for paternity testing.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print







