





INTRODUCTION
Heat stress (HS) can directly endanger the health status of pigs in addition to the reduced feed intake [1]. The two major insults that result from HS conditions are oxidative stress and respiratory alkalosis which both comprehensively compromise pig health. Pigs redistribute blood flow to peripheral tissues [2] and increase their respiration rate to maximize heat dissipation [3] in HS conditions. Consequently, the reduced blood flow in the splanchnic tissues along with an elevated body temperature triggers oxidative stress in the intestine [4,5] and skeletal muscle [6] of pigs. Oxidative stress, a result from imbalanced free radical and antioxidant capacity, damages lipid, proteins and nucleotides thereby expansively and negatively affecting the functionality of cells and organs [7]. Particularly, the increase of oxidized proteins is associated with various metabolic disorder in livestock [8,9]. On the other hand, hyperventilation reduces blood CO2 and consequently triggers alkalosis. Respiratory alkalosis can disturb various biochemical and physiological process of the body such as decreasing calcium ionization [10] and increasing lactate production [11]. Therefore, alleviating the oxidative stress and respiratory alkalosis is crucial for maintaining pig health and production, and nutritional supplementations may provide a flexible and economical strategy for mitigation.
Vitamin E (VE) is a lipid soluble antioxidant which neutralizes free radicals. The National Research Council recommends 11 IU/kg VE for growing pigs raised in normal conditions [12]. However, it remains unknown if pigs require higher amount of dietary VE for counteracting HS-induced oxidative stress. A recent study from our research group has demonstrated that a 14-d supranutritional antioxidant supplementation (100 IU/kg VE plus 1.2 ppm selenium) could not only mitigate oxidative stress, but also effectively ameliorate the respiration rate and respiratory alkalosis in heat-stressed sheep [13], suggesting that a short-term supranutritional antioxidant supplementation effectively mitigate the HS-induced oxidative stress and exhibit a novel function in maintaining acid-base balance. Therefore, the objective of the present study was to investigate the effects of supranutritional VE on redox status, and to confirm the novel function of VE in ameliorating respiratory alkalosis in the heat-stressed pigs. It is hypothesized that a short-term supplementation with a supranutritional amount of VE (200 IU/kg) before and during a heat event may mitigate oxidative stress and respiratory alkalosis in pigs subjected to HS.
MATERIALS AND METHODS
The experimental protocol was approved by the Animal Ethics Committee of Faculty of Veterinary and Agriculture Sciences at University of Melbourne.
Animals and experimental design
The experiment followed a 2├Ś2 factorial design which included two dietary treatments and two temperature conditions. Twenty-four growing female pigs (Large White├ŚLandrace, 24┬▒2 kg, mean┬▒standard deviation) were randomly assigned to a control dietary treatment which was formulated to meet or exceed NRC standards [12] and contained 17 IU/kg VE (Control) or a high VE diet (HiVE) which was fortified with 0.075% VE premix containing synthetic ╬▒-tocopherol acetate (Rovimix E50, DSM Nutritional Products, Basel, Switzerland) to increase VE concentration to 200 IU/kg (Table 1). After a 14-d dietary supplementation, six pigs from each dietary treatment were housed in either thermoneutral (TN; 20┬░C, 45% relative humidity) or cyclic HS conditions (35┬░C, 09:00 h to 17:00 h; 28┬░C, 17:00 h to 09:00 h, 35% to 45% relative humidity) for 7 d. Pigs were housed individually in metabolic crates with metal mesh floor during the experiment. To minimize the confounding effects of different feed intake between TN and HS groups, the feed allowance was restricted to approximately 75% of voluntary feed intake of the pigs housed under TN conditions. The feed allowance was predicted from anticipated voluntary feed intake of heat-stressed pigs [3]. The pigs were fed at 09:00 h and 17:00 h daily, and the feed refusals were weighed at 09:00 h daily to calculate individual feed intake. Pigs were allowed free access to water via nipple drinkers.
Physiological monitoring
All pigs were monitored for physiological signs of HS including respiration rate and rectal temperature three times daily at 09:00 h, 13:00 h, and 16:00 h during the 7-d thermal exposure period. Rectal temperature was measured using an electronic thermometer (Fast-Read, Livingstone Pty Ltd., Rosebery, NSW, Australia). The respiration rate was visually counted within 20 s using a stop watch. As a precaution, pigs were evacuated from the HS room if the rectal temperature exceeded 41┬░C until their rectal temperature dropped below 40┬░C. One pig was evacuated for 1 h due to hyperthermia and was returned to the HS room without incident.
Blood collection and blood gas detection
Blood samples were collected via venipuncture from the jugular vein at 09:00 h and 15:00 h on d 7 from the start of the thermal exposure, and the blood samples were collected into a non-anticoagulant, a heparinized and an ethylenediaminetetraacetic acid (EDTA) coated vacutainers (BD, North Ryde, NSW, Australia). A sample of fresh blood (0.2 mL) from each non-anticoagulant vacutainer tube was immediately injected into a blood gas analyzer (EPOC, Alere, Waltham, MA, USA) for measuring CO2 partial pressure (pCO2), O2 partial pressure (pO2), bicarbonate concentration and, blood pH. The blood samples in the heparinized and EDTA coated tubes were centrifuged for 10 min (2,000 ├Śg, 4┬░C). Approximately 0.5 mL of leukocytes from each EDTA coated vacutainer tubes were harvested and added into 1 mL RNAlater solution (Lifetechnologies Pty Ltd., Mulgrave, VIC, Australia), then the leukocyte samples were stored at 4┬░C overnight before stored at ŌłÆ20┬░C. Heparinized plasma samples were stored at ŌłÆ80┬░C till further analysis for oxidative stress biomarkers.
Oxidative stress biomarkers
Derived reactive oxygen metabolites (ROM), biological antioxidant potential (BAP), and thiol groups (SHp) in the heparinized plasma were measured using an integrated analytical system (FREE Carpe Diem, Diacron International, Grosseto, Italy). The ROM assay measures the blood concentration of hydroperoxides, a class of chemical oxidant species belonging to the wider group of ROM, its values were expressed as H2O2 (mg/dL) equivalency. The ROM assay has been validated and used in pigs for estimating H2O2 concentration [14,15]. The BAP test provides a combined measurement of several antioxidants including uric acid, ascorbic acid, proteins, ╬▒-tocopherol, and bilirubin by measuring the capacity of the plasma sample to reduce ferric (Fe3+) to ferrous (Fe2+) iron. The results of BAP are expressed in mmoles per liter of reduced iron. The SHp assay measures the antioxidants contains thiol group such as glutathione and lipoic acid. Concentration of advanced oxidized protein products (AOPP) was measured by method described previously [16], and the values were expressed as ╬╝M of chloramine T equivalents.
Detection of mRNA expression
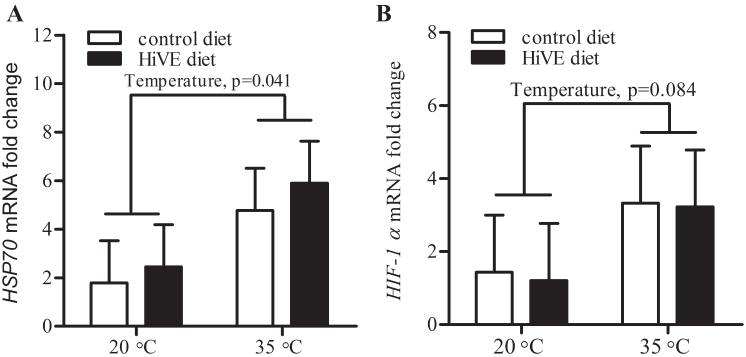
Leukocytes were pelleted and separated from the RNA preservative after 1 min centrifugation at 1,000 g at room temperature. Total RNA in the leukocytes was extracted in the acid-phenol solution according to instructions of the manufacturer (RiboPure, Lifetechnologies Pty Ltd., Australia). The RNA concentration and quality were evaluated by a RNA analysis system (Experion, Bio-Rad Laboratories, Inc., Hercules, CA, USA). Then 800 ng RNA of each sample in triplicates was transcribed to cDNA using reverse transcriptase (Superscript III, Lifetechnologies Pty Ltd., Australia). The synthesized cDNA samples were stored at ŌłÆ20┬░C. Primers sets for swine 18S ribosome RNA (r18s), hypoxia induced factor-1╬▒ (HIF-1╬▒) and heat shock protein 70 (HSP70), were referenced from previous work [17]. Twenty-five ╬╝L reactions were prepared according to the manufacturerŌĆÖs instruction using SYBR green supermix (Lifetechnologies Pty Ltd., Australia), and 100 nM of each forward and reverse primer were added in each reaction. Each sample was amplified in triplicates and fluorescence of SYBR green was quantified in iQ5 Real Time PCR Detection System (Bio-Rad Laboratories Inc., USA). Each polymerase chain reaction (PCR) plate included five serial dilutions of a pooled cDNA as a standard curve, a non-template negative control, and a blank control to calculate the PCR efficiency. The PCR performed after denaturing the cDNA samples at 95┬░C for 10 min, then 45 cycles consisting of three stages were applied. A protocol (95┬░C for 30 s, 60┬░C for 30 s, and 72┬░C for 30 s) was used for r18s and HSP70, the other protocol (95┬░C for 30 s, 55┬░C for 30 s, and 72┬░C for 30 s) was used for HIF-1╬▒. A melting curve was included to verify the single PCR amplification. The abundances of mRNA were calculated as fold changes.
Statistical analysis
Data were analysed by general linear model with Repeated Measures in SPSS Version 24 (IBM SPSS, Chicago, IL, USA). Each individual pig was treated as an experimental unit. For rectal temperature and respiration rate analysis, days of thermal exposure (d 1 to d 7) and time of the day (09:00 h, 13:00 h and 16:00 h) were set as within-subject factors, and environmental temperature and dietary treatment were set as between-subjects factors. The results of rectal temperature and respiration rate were pooled across ŌĆ£dayŌĆØ factor and presented as means and standard errors of means (SEM) of temperature├Śdiet├Śtime. For the blood variables, the time of blood sampling (09:00 h and 15:00 h) were set as a within-subject factor, and environmental temperature and dietary treatment were set as between-subjects factors. The results of blood variables were pooled across ŌĆ£timeŌĆØ factor and presented as the means and SEM of temperature├Śdiet. The difference among the treatments was considered to be significant if pŌēż0.05, and a trend was considered if pŌēż0.10.
RESULTS
Feed intake and physiology
Restricting the feed allowance ensured that the average daily feed intake (ADFI) averaged 1.46 kg┬▒0.002 (mean┬▒SEM) across all the treatment groups. Average daily feed intake was not affected by temperature (p = 0.82) or diet (p = 0.81; data not shown).
Heat stress increased rectal temperature (38.3┬░C vs 39.6┬░C for TN and HS, p<0.001; Figure 1A), and there was an interaction between temperature and time (p<0.001) such that the rectal temperature increased from 38.6┬░C to 39.9┬░C from 09:00 h to 13:00 h, and maintained at 40.1┬░C until 16:00 under HS conditions, whereas the rectal temperature was constant across the day for pigs under TN conditions. The HiVE diet did not affect rectal temperature (p = 0.78), and the interaction between diet and temperature or time was not significant.
Heat stress greatly increased respiration rate (31 to 124 breaths/min for TN and HS, p<0.001; Figure 1B) and there was an interaction between temperature and time (p<0.001) such that the respiration rate increased from 51 to 154 breaths/min between 09:00 h and 13:00 h before plateauing at 166 breaths/min until 16:00 h, whereas the respiration rate was constant across the day under TN conditions. The HiVE diet did not affect respiration rate (p = 0.99), and the interaction between diet and temperature or time was not significant.
Blood gas variables
Blood pCO2 decreased (p = 0.002) from 58.7 to 46.8 mm Hg and bicarbonate decreased (p = 0.002) from 37.0 to 33.1 mM during HS, whereas pH increased (p = 0.012) from 7.42 to 7.46 during HS. Blood pO2 was not affected by HS. While the main effects of HiVE on these variables were not significant, there were interactions between temperature├Śdiet (all p<0.05) on blood, bicarbonate and pH, indicating that the blood gas variables of the pigs fed control and HiVE diet were affected differently by thermal conditions (Table 2). Specifically, compared with the pigs fed control diet, the pigs fed HiVE diet had a similar blood pCO2, bicarbonate and pH in TN condition but a greater level of pCO2, a greater level of bicarbonate and a lower blood pH in HS condition.
Oxidative stress markers and gene expression
Heat stress markedly increased mRNA abundance of HSP70 (1.5 vs 5.4, p = 0.041) and tended to increase the mRNA abundance of HIF-1╬▒ (1.3 vs 3.3, p = 0.084) in the leukocytes. The mRNA abundance of HSP70 or HIF-1╬▒ was not affected by HiVE diet (Figure 2A, 2B). Neither HS or HiVE diet had a main effect on ROM concentrations; however, there was an interaction between temperature and diet (p = 0.047) such that the pigs fed HiVE diet had lower ROM concentrations than the pigs fed control diet in TN conditions but became similar as control group in HS conditions (Table 3). Heat stress reduced plasma BAP from 2.97 to 2.73 mmol/L (p = 0.003), whereas HiVE diet did not affect plasma BAP. Heat stress tended to increase AOPP concentrations from 16.4 to 26.9 ╬╝mol/L (p = 0.067), whereas HiVE diet did not affect plasma AOPP concentrations. Neither HS nor dietary VE supplementation affected plasma SHp concentrations.
DISCUSSION
The hypothesis of this study was that a short-term supranutritional amount of VE (200 IU/kg) supplementation may mitigate impacts of HS on physiological responses, respiratory alkalosis and oxidative stress in pigs. Our data indicated that the VE supplementation used in the present study did not mitigate oxidative stress in heat-stressed pigs, but partially alleviated HS-induced respiratory alkalosis.
Heat stress increased rectal temperature by 1.3┬░C, while respira tion rate increased three-fold to aid heat dissipation. The increased respiration rate of HS pigs caused a 20% decline in blood pCO2 which presumably reduced the formation of carbonic acid that is the driving factor for the increased blood pH. As a metabolic compensatory mechanism to maintain blood pH, blood bicarbonate which is the main base contributor, was reduced by 12% in the current study. Despite this, a normal blood pH could not be maintained, as the blood pH was increased by HS from 7.42 to 7.46. The whole process was a typical hyperventilation-caused respiratory alkalosis which could not be compensated by metabolic acidosis. Similarly, respiratory alkalosis occurs in sheep [13], poultry [18] and pigs [19] subjected to HS conditions. In agreement with the finding in the antioxidant supplemented sheep during HS [13], the HiVE diet ameliorated the loss of blood pCO2 by 50%, which prevented the increase of blood pH and spared the reduction of blood bicarbonate concentration. Although the mechanisms by which HiVE ameliorated the loss of blood pCO2 are not known, the effect was independent of the amelioration of hyperventilation which was observed in the heat-stressed sheep, because HiVE diet did not reduce respiration rate of the heat-stressed pigs in present study. In other words, dietary HiVE may have had a direct effect in preserving blood CO2 concentration resulting in mitigation of the HS-induced respiratory alkalosis. In agreement with previous observations, blood oxygen concentration were unaffected by HS [4,17], but leukocyte HIF-1╬▒ mRNA abundance tended to increase in the heat-stressed pigs, which is possibly due to the activation by oxidative stress [20,21].
Heat stress triggered oxidative stress as evidenced by the in creased AOPP in plasma. The increased oxidative stress was possibly due to the compromised antioxidant defense system, because BAP, the measurement of overall antioxidant capacity of plasma, was reduced by 9% during HS. Moreover, the reduction in BAP is consistent with observations made in ruminants subjected to HS conditions [13]. The increased oxidative stress is associated with the occurrence of hyperthermia in the heat-stressed pigs, as evidenced by the increased mRNA abundance of HSP70 in leukocytes. However, unlike in sheep subjected to a similar magnitude of HS [13,22], plasma ROMs concentrations were not affected by HS conditions, suggesting that reduction of BAP was not caused by increased free radicals. Other factors which affect the antioxidant stability or regeneration should be considered.
Dietary VE supplementation did not alleviate oxidative stress in the pigs subjected to HS, because neither the decreased BAP nor the increased AOPP was alleviated by the HiVE diet. The total duration of VE supplementation was 3 wk in present study which included the 2 wk before and 1 wk during the thermal exposure period. The 3-wk supplementation should have been sufficient to enhance tissue stores of VE. For example, the plasma and muscular VE reached 80% of the maximum concentration in pigs after a 2-wk supplementation with VE (35 IU/kg vs 300 IU/kg) [23]. Indeed, the HiVE diet reduced blood ROMs concentrations by 20% in the pigs housed under TN conditions, suggesting the 200 IU/kg VE is superior to the dietary VE concentration recommended by NRC [12] in neutralizing free radicals when pigs are in normal physiological status. However, the difference of ROM concentration between the pigs fed HiVE and control diet diminished after exposing to HS conditions, which was possibly due to the alteration in free radical production by HS although the exact mechanism remains unknown. We speculate that the contribution of VE to the alleviation of oxidative stress in heat-stressed pigs may be limited by the absence of synergisms of VE with other antioxidants such as VC and glutathione, because the regeneration of VE requires the participation of VC and glutathione peroxidase [24]. A previous study conducted by our research group has shown that combination of VE (100 IU/kg) and Se (1.2 ppm) successfully alleviated the reduction of BAP and the increase of AOPP in the sheep subjected to similar level of HS [22]. Similarly, in broilers supplemented with a combination of VE (125 mg/kg) and Se (0.5 ppm) had reduced plasma malondialdehyde concentration than the broiler separately supplemented VE or Se under hot conditions [25]. Moreover, a combination of supranutritional VE (200 IU/kg) and Se (1.0 ppm) reduced the oxidative stress markers in the intestine of pigs subjected to a 2-d HS [17]. Future experiments should investigate the combinations of dietary antioxidants in reducing the oxidative stress in heat-stressed pigs.
In summary, a 3-wk supplementation with supranutritional amount of VE (200 IU/kg) did not alleviate oxidative stress in growing pigs subjected to a 7-day cyclic HS. However, dietary VE supplementation did prevent respiratory alkalosis observed during HS conditions and may be an effective dietary supplement for use in a summer ration.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







