




INTRODUCTION
Animal behavior occurs as a response to external environment stimulation and internal motivation and generally appears to favor survival. Behavioral observation is an efficient way to quickly and precisely understand animals’ internal/external status, and is also a definitive method for assessing animal-centered welfare status [1]. For this reason, behavioral study on livestock including cattle has been actively performed around the globe [2–7]. Recent outbreaks of foot-and-mouth disease and avian influenza in Korea highlighted weaknesses in intensive animal husbandry methods. Consumers are paying more attention to the breeding environment of animals and the production of livestock commodities. As a consequence, there is increasing demand for an ecofriendly animal production system that considers animal welfare. Hanwoo (Bos taurus coreanae) are cattle native to Korea. Over the course of 4,000 years [8], they adapted to Korean weather, which has four distinct seasons and ranges from hot and humid in the summer to cold and dry in the winter. The cattle are generally raised in cow houses because more than 70% of Korean territory is mountainous, and they are fattened for more than 30 months to increase the marbling score of their meat. Despite the unique genetic and environmental conditions of Hanwoo, behavioral research based on these factors is extremely limited. Research on cattle behavior that focuses on Hanwoo can be divided into: first, analysis of livestock behavioral repertory according to the genetic factors, and natural environment such as species, growth stage, gender, physiological state, environmental temperature, and humidity [9–11]; second, behavioral changes due to differences in rearing system and management methods [12–16]; third, studies on behavioral response of cattle according to handling methods of livestock; and fourth, analysis methods of behavior [17]. In particular, the first subject of these is the most important and becomes the basis of all behavioral studies since it can provide the fundamental data of behavior relating to genetic and basic environment of the cattle. However, basic researches on Hanwoo’s behavior are very rare and existing studies are limited in subject and scope. Therefore, comparing results between references is not easy, and evaluating animal welfare for Hanwoo is difficult. Like other animals, cattle have species-specific behavior patterns, and the frequency of the behavioral manifestation and differences over time can vary according to the genetic background and environmental conditions, including breeding method. For mutual development of the rearing method and animal welfare, fully understanding the relevant livestock must precede, because relevant alternatives can be found only after correctly comprehending the physiological and emotional states of the livestock. Thus, this study will investigate behavioral characteristics of Hanwoo steer based on seasons and growth stages to collect baseline data to enable measurement of animal welfare.
MATERIALS AND METHODS
Animals and housing
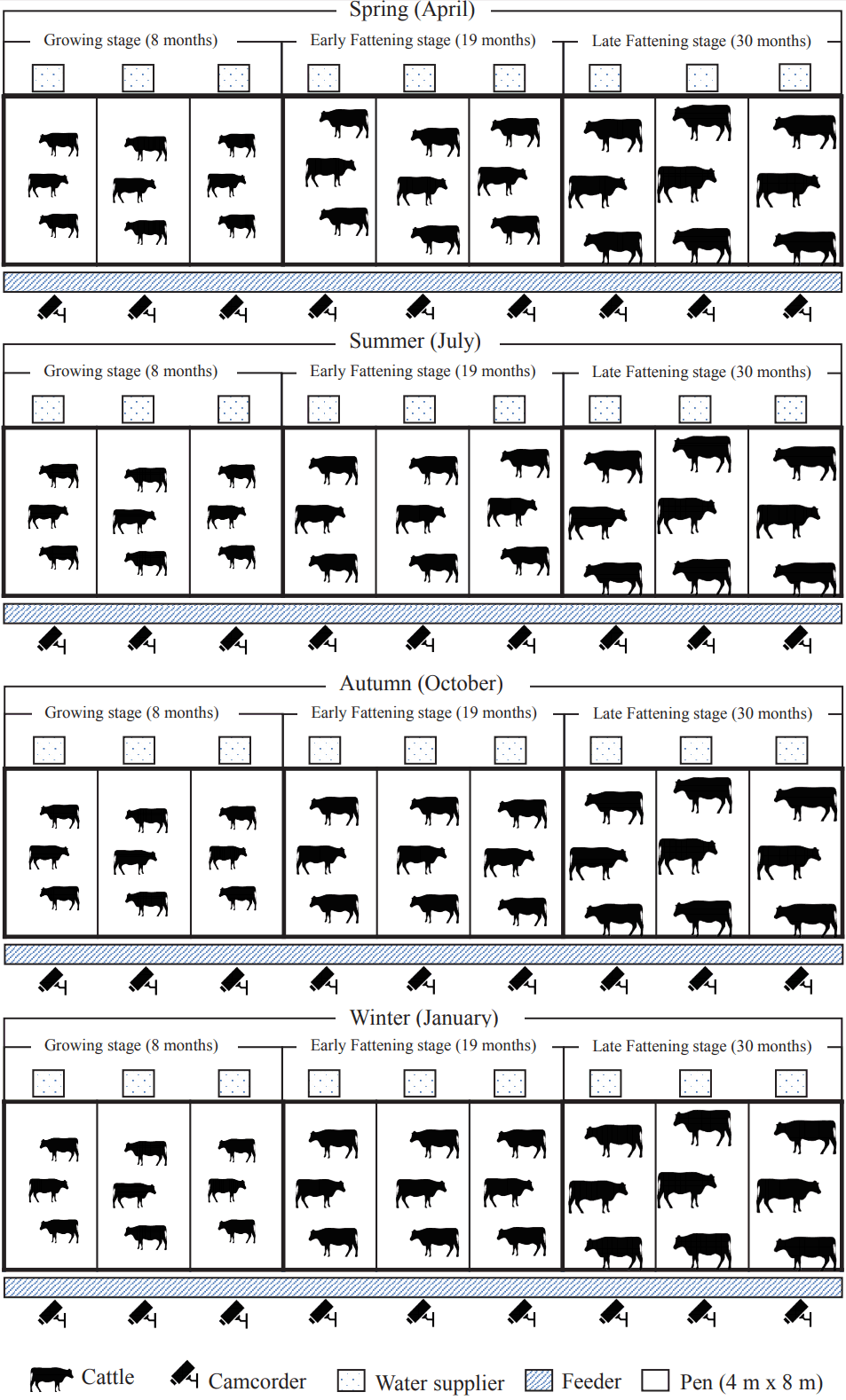
This experiment was conducted under the approval of the Institutional Animal Care and Use Committee of Konkuk University (IACUC No. KU 12097). The experiment was performed at a beef cattle farm (farm Kodaemi, South Korea) from 30 January to 14 October 2013. The Information on experimental design is presented in Figure 1. To perform the experiment, this research experimented 108 Hanwoo steers, introduced from calf auction markets and gelded in their 6 months. In order to observe their behavior, steers were planned to be divided 27 heads respectively in Spring (April), Summer (July), Autumn (October), and Winter (January), based on four seasons. Subsequently, three growth stages for each season were divided into: growing stage (GS, 8 months), early fattening stage (EFS, 19 months), and late fattening stage (LFS, 30 months), and nine steers were assigned to each growth stage ([9 heads×3 growth stages]×4 seasons = 108 heads). Body weights were measured three days before the start of the experiment, the mean weights were 236±27 kg in the GS, 507 kg±31 in the EFS, and 753±52 kg in the LFS. In consideration of proper individual space of cattle [15] steers were housed at all times in pens that provided 10.67 m2 per head, and each pen accommodated three animals in the same growth stage. Feed was mixed in the form of total mixed ration, which was based on the standard diet for Hanwoo. The chemical composition of the diet is presented in Table 1. The animals had ad libitum access to feed and water. Sawdust was spread on the floor to a thickness of 15 cm, and it was replaced regularly to maintain dry conditions for the animals. The steer house was covered with a 4.5-m roof that facilitated ventilation and included a winch curtain, which blocked the wind from entering in the winter. Otherwise, the curtain was opened most of the time.
Behavioral observations and analysis
Aiming to analyze the expression pattern of daily behavior of Hanwoo steers along with the season and growth stage, nine closed-circuit television cameras connected to a multiplexer were installed in front of each pen a height of 2.2 meters to record the animals’ behavior. All behaviors of steers were continuously recorded during 13 day-time hours (06:00 to 19:00 h) with three replications. Recorded behaviors were collected on three days each season. Seasonal ambient temperatures, relative humidity, temperature-humidity index (THI) [18] for the observation dates were measured five times a daytime (07:00 to19:00 h), and the range of data was 4°C to 24°C/16% to 77%/42 to 67 in the spring; 25°C to 35°C/54% to 90%/76 to 86 in the summer; 13°C to 26°C/ 42% to 89%/56 to 72 in the autumn; and −9°C to 2°C/15% to 63%/24 to 46 in the winter (Table 2).
Twelve behaviors of steers were observed along with the season and growth stage. Categories of behavior included standing, lying, eating, ruminating, standing-resting (s-resting), lying-resting (l-resting), walking, drinking, sleeping, self-grooming, social grooming, and sniffing (for definitions, Table 3). Expressed behaviors were recorded in terms of a unit of time (minute or second) or frequency. In addition, the mean bout frequencies of standing, lying, and eating were measured. A single bout refers to the period from the start of a behavior to its end, and the total number of bouts was designated as the bout frequency. Standing (or lying) bout frequency referred to the total number of standing (or lying) behaviors by individuals during the observation period; that is, one bout meant a sitting or standing individual stood up or lay down and then lay down or stood up again. Eating bout frequency referred to the total number of eating behaviors of an individual, with one eating bout meaning that an individual took feed into its mouth from the feed tray and chewed it for no longer than 20 consecutive seconds. Despite distinct differences between groups regarding the mean bout durations for standing, lying, and eating, large standard deviations were produced because sessions of longer times and shorter times were separated. Therefore, the duration of each bout was analyzed differently. First, the total time spent in standing, lying, and eating behaviors was examined, and then the duration was investigated to set duration points (DPs; standing, 60 min; lying, 30 min; eating, 10 min) for bouts of each behavior. Based on the DPs, each behavioral bout duration was divided based on whether it was long or short, and the distribution rate (%) was calculated for the total frequency of the two sessions of the groups. If the above DP frequency ratio was higher, the bout duration could possibly be longer, and if the below DP frequency ratio was increased, the bout duration could be shorter.
Statistical analysis
The mean time and frequency values for 13 behaviors of cattle during 13-hour time periods were compared among the four seasons and the three growth stages. The mean bout frequencies of standing, lying, and eating were also compared among the four seasons and the three growth stages. Factors were analyzed by using analysis of variance or the general linear model procedure in SAS (ver. 9.1, SAS Institute, Cary, NC, USA). When a statistically significant difference was identified, Duncan’s multiple range test was used to detect statistical significance (p<0.05) among treatment groups.
RESULTS
Table 4 shows the average time or frequency of 13 behaviors of steers according to season and growth stage. Daytime standing times were significantly the longest in the summer (p<0.05) at all growth stages (GS 749.89 min, EFS 659.89 min, LFS 614.11 min). The cattle in the LFS group showed higher significance even in autumn as well as in summer than other seasons (p<0.05). Standing times were second longest for all growth stage groups in the autumn. In the spring and summer, GS (536.44 and 749.89 min, respectively) standing time was significantly longer than in the EFS group (455.11 and 659.89 min, respectively) and the LFS group (482.22 and 614.11 min, respectively) (p<0.05). In contrast, standing bout frequency (Table 5) for the GS group was significantly lower in summer (2.89) than in other seasons (p< 0.05); the same pattern was seen in the EFS and LFS groups. In Table 6, above DP frequency distribution for summer standing was 62% in the GS group, 73% for the EFS group, and 73% for the LFS group, far higher than in the other three seasons.
In winter and spring, lying times were the highest for all growth stage groups, and the GS group had significantly longer times in the winter (319.06 min) than in the other three seasons. The EFS group showed significantly longer times (324.89, 120.11, 311.11 min) in the spring, autumn, and winter than in the summer. The LFS group showed significantly longer times (290.89, 297.00 min) in the spring and winter than in the summer and autumn (p< 0.05). Excluding spring values for the EFS group, average lying times were progressively shorter in the winter, spring, autumn, and summer. Summer lying times (GS, 30.11 min; EFS, 120.11 min; LFS, 152.78 min) were significantly the shortest at all growth stages (p<0.05). Among the groups, the GS group had shorter summer lying times than the other two stages and was significantly different from the LFS group (p<0.05). Meanwhile lying bout frequency (Table 5) was the lowest in the GS and EFS groups in the summer. The GS group (2.78) showed lower levels in the summer than in the autumn and winter, and the EFS group (3.56) showed significantly lower levels in the summer than in any other season (p<0.05). Below DP frequency distribution for summer lying in the GS group was 96%; EFS, 50%; and LFS, 58% (Table 6).
The GS and EFS eating times in the summer (GS, 151.39 min; EFS, 143.23 min) and winter (GS, 177.17 min; EFS, 142.73 min) were shorter than those in the spring (GS, 246.50 min; EFS, 188.29 min) and autumn (GS, 268.18 min; EFS, 216.09 min) (p<0.05). However in the LFS group, eating times showed no seasonal difference. GS eating times in spring, autumn, and winter were significantly longer than the other groups (excluding LFS in winter) (p<0.05). For eating bout frequency (Table 5), the GS group showed significantly higher values in the summer than in the other three seasons (p<0.05). Although no significance difference was found, EFS eating bout frequency was also highest in the summer compared with the other seasons. However, below DP frequency distribution for summer eating was 91% in the GS group; 81% in the EFS group; and 84% in the LFS group, which were relatively higher levels (Table 6).
Ruminating time in the LFS group (spring, 104.06 min; sum mer, 117.87 min; autumn, 89.44 min; winter, 111.64 min) was significantly lower than those in the other growth stage groups in all seasons (p<0.05).
Standing-resting times were the longest in the summer for the GS group (414.65 min) and the EFS group (406.81 min), while the LFS group had their longest S-resting times in the summer (376.87 min) and autumn (386.70 min) (p<0.05). The LFS s-resting times were longer in the spring (297.99 min) and in autumn (386.70 min) than those for the other groups (p<0.05).
Lying-resting time was significantly higher in the winter for the GS group (204.08 min), in spring for the EFS group (224.77 min), and in spring (247.14 min) and winter (220.25 min) for the LFS group (p<0.05) than in other seasons, with times becoming higher as the ambient temperature rose. The GS l-resting times in the spring (149.56 min) and in the summer (17.85 min) were significantly shorter than for the other growth stage groups (p< 0.05). This result occurred because of the shorter lying time than in the other groups. L-resting times in most seasons tended to be longer at older growth stages.
The GS (38.67 min) and EFS (16.62 min) walking times were the longest in the summer among all seasons (p<0.05). Although not significant, walking time in the summer was also longest for the LFS group. In spring (6.70 min) and summer (38.67 min), GS walking times were significantly the longest among all growth stage groups, excluding the spring value for the LFS group (5.37 min) (p<0.05).
The GS (39.38 min) and LFS (20.78 min) drinking times were longer in summer than in the other seasons, and in summer, the GS group had the longest times among all growth stages (p<0.05).
Sleeping time was significantly longer in the spring (29.20 min) and winter (33.25 min) for the GS group, and the EFS group had a significantly longer sleeping time in the spring (33.85 min) than in other seasons. The GS and EFS groups recorded significantly a shorter sleeping time in the summer (0.84 min and 0.90 min, respectively) than in the other seasons (p<0.05). The LFS group, however, showed no significant seasonal differences. The LFS group recorded significantly longer times in the summer (19.69 min) and the autumn (27.19 min) than cattle in the other groups (p<0.05). This finding indicates that LFS cattle, unlike GS and EFS cattle, did not reduce their sleep behavior even in warm conditions.
Self-grooming time was significantly the highest for all growth stage groups in the winter (GS, 668.22 s; EFS, 1,838.75 s; LFS, 312.86 s) than in any other season. The LFS self-grooming times in spring (156.11 s) and winter (312.86 s) were the lowest among all groups (p<0.05).
Social-grooming times in the EFS and LFS groups were the highest in the spring among all seasons and the lowest in the autumn (p<0.05). In the spring and winter, social-grooming times in the EFS group (spring, 317.13 s; winter, 231.56 s) were the highest, while the LFS group had the lowest (spring, 175.00 s; winter, 66.56 s) (p<0.05).
The GS and EFS sniffing frequencies were highest in the spring (GS, 20.67; EFS, 29.56), while the highest values occurred for the LFS group in the spring (17.44) and winter (19.38) (p<0.05). In particular, LFS sniffing frequency in the winter was significantly higher than GS sniffing frequency (8.67) (p<0.05). Sniffing mostly occurred when choosing a place to sit down leading to lying activities.
Generally, in the GS group, i) standing, eating, ruminating, walking, and drinking behavior times were longer, ii) while lying, s-resting, and l-resting times were shorter. In the LFS group, i) s-resting, l-resting, summer lying, and summer and autumn sleeping times were longer, ii) whereas self-grooming, social-grooming, and summer walking times were shortened, showing more dynamic movement declined as cattle grew. In general, as ambient temperature rose, i) cattle standing, s-resting, walking, and drinking behaviors increased, ii) while lying, l-resting, and eating behaviors decreased. Along with this, the characteristic of behavioral pattern should be considered, where iii) time spent standing and standing duration per bout increased while standing bout frequency decreased, iv) time spent lying, lying bout frequency, and lying duration per bout all decreased, also, v) eating bout frequency increased while time spent eating and eating duration per bout decreased. In contrast, as ambient temperature dropped, i) lying and self-grooming behaviors increased, ii) but standing behaviors were reduced.
DISCUSSION
Standing times among cattle tend to be longer as the ambient temperature increases. Ha et al [9] displayed a completely opposite result from this research, regarding that total time spent standing of steers increased as the weather gets colder which was reported because the steers evaded standing due to high temperature stress of summer. However, Allen et al [4] reported cow standing behavior was affected once THI reached 68. Given this finding, it is deemed that since the summer THI assessed in our research was 75 to 85.6 (Table 2), steer standing time was strongly affected by heat condition. Igono et al [19] and Zahner et al [20] found increased ambient temperature increases standing times in heat-stressed cattle. Anderson et al [21] reported that an increase in core body temperature (CBT) may be positively correlated to the amount of time that cows stand. Allen et al [4] found that cows stand up as CBT increases, and they also found that a CBT of 38.93°C was associated with a 50% likelihood that a cow would be standing; therefore, standing may help cool cows. Although these studies investigated different cattle breeds under different conditions, their results were consistent with our findings, thus we confirmed that standing is an important behavior in assessing cattle’s heat stress. Due to the high temperature and humidity during summer in South Korea, the overall level of THI was extremely high. Therefore, increase of time spent standing can be recognized as an important change in detecting the level of heat stress experienced by Hanwoo during hot and humid summer. However, the other possibility of another factor related to the standing of cattle should be considered through the research of Ha et al [9], who reported completely opposite result from this research. Accordingly, in order to utilize standing as the behavioral index considering the original gene and environment of Hanwoo, more accurate parameter research should be conducted additionally related to other factors such as THI, CBT, bedding type and state, etc.
Summer standing time became shorter as cattle moved from the GS to the LFS. Younger beef cattle were found to be more sensitive to heat. Although the total standing time in summer was longer than any other season, the standing bout frequency being significantly less than the other seasons meant that the average standing duration per bout was also long (time spent of behavior = bouts×duration per bout). This conclusion is supported by the distribution of above DP frequency of summer standing being greater. Allen et al [4] reported that the duration of standing per bout lasted longer when a cow had a greater CBT, which is similar to our finding. Therefore, the summer standing behavior pattern of steer had the lowest frequency, although its duration was the longest and the total standing time was the longest among all seasons.
Average lying time increased at lower ambient temperatures. Graunke et al [22] observed beef cattle behaviors in Scandinavia and found higher lying time was associated with precipitation such rain and snow. Our research found no large difference in Hanwoos’ lying time between the spring and winter, showing that the cattle were successfully adapted to the winter cold. However, the GS group was found to have a significantly longer lying time in the winter than in the spring, thus more care seems necessary for GS cattle in the winter. The time spend lying down is an important measure of cattle comfort [23]. Cattle can only sleep in a comfortable lying down position, and resting in this position is more effective for regaining physical strength than resting in a standing position [1]. Reducing the time that cattle can lie down results in both behavioral and physiological evidence of stress [24]. The EFSA [25] and Lee et al [26] found that cows spend an average of 12 h/d lying down. A reduction in lying time also means an increase in standing time. It seems necessary to work to address the rapid reduction in summer lying time and ensure a certain level of lying time for cattle to improve animal welfare. In summer, lying bout frequency was the lowest, and below DP frequency distribution of summer lying was greater at all growth stages. Kanjanapruthipong et al [27] reported that the duration of lying bouts decreased linearly with increasing THI, which was similar to the finding in our study. Lying behavior of beef cattle (GS, in particular) was found to be shorter or lower in terms of time, bouts, and bout duration in the summer compared with the other seasons. In contrast, above DP frequency distribution of winter lying was 73% in the GS group, 77% in the EFS group, and 68% in the LFS group, indicating that lying time and bout duration are remarkably longer in the winter, than in the other seasons. Consequentially, heat condition changes were found to possibly cause not only quantitative change in cattle lying behavior but also general changes in lying behavior patterns.
The eating time was short in the summer and the winter. Generally, the appropriate temperature for cattle ranges from 0°C to 20°C as reported by Won [28]. The ambient temperatures of summer (25°C to 35°C) and winter (−9°C to 2°C) in this study were not appropriate heat conditions for the cattle. Therefore, the stress from heat and cold was deemed to lead to shorter eating times. However, cattle are inclined to have replenishment eating after sunset when the summer environmental temperature goes down [1], so that time spent eating per day could somewhat increase considering the possibility of replenishment eating. In addition, the eating time in the GS group was longer than in the other growth stage groups. Mimura et al [1] reported that cattle eating time was shorter as total digestible nutrition (TDN) increased, and it was no longer than 4 hours a day if the cattle were mostly fed grains. Among the experiment cattle, TDN (based on dry matter) increased from GS (70.06%), to EFS (74.26%), and to LFS (81.27%) (Table 1), but eating time because shorter in contrast as cattle grew from GS and into LFS. In the present study, eating time was found to be approximately 3 hours or less, given that this was recorded during 13 day-time hours, the results are estimated to be almost similar to those of the referred studies of Mimura et al [1], Cozzi et al [29], Mazzenga [30]. The eating bout frequency was highest and the distribution of below DP frequency of eating was higher in the summer, which mirrors the results of [27]: “The duration of eating bouts decreased linearly with increasing THI”. Consequently, eating frequency was highest in summer for GS and EFS cattle, but eating time was mostly short; therefore, compared with the other seasons, the shortest total eating time occurred. As a result, the summer eating behavior was characterized by short, frequent bouts of eating, which was most clearly revealed in the GS group.
Regarding the ruminating time, the LFS group had shorter times than the other groups. Ruminating time increases as the intake amount and neutral detergent fiber (NDF) content rise, but decreases as crude protein (CP) content rises and feed particles become smaller [1]. In our research, CP, crude fiber, acid detergent fiber, and NDF were found to decrease as cattle grew (Table 1). Therefore, it’s considered that time spent ruminating in LFS group significantly decreased. Also, time spent ruminating of this research showed the result of 13 hours in the day time, Mimura et al [1] reported that over 60% of time spent ruminating of cattle was taken while lying posture during night time, and Cozzi et al [29] reported that 75% (249 min) out of daily average time spent ruminating (332 min) of experimental cattle was spent between the sunset and the dawn. When aggregating the result of beef cattle, consuming hours during the day time appeared similar in this research to other data.
Mimura et al [1] reported that cattle tend to gather together in warm seasons to minimize attacks from flies or blood-sucking insects through minimizing their body surface areas. The increased walking time in this research seemed mainly for this reason. During most of the observation time, the cattle kept shaking their tails and ears, and if one individual moved, the rest of the group followed, consequentially increasing their walking time. Such behaviors support the assessment. Based on the walking time of the GS group being longest, it could be concluded that the younger growth stage is associated with longer walking time in the hot condition.
Since self-grooming was found to increase in winter and spring where external parasites such as tick, flea, mite, lice, demodex etc., increase inside the fur of the body, longer self-grooming times in winter and spring are viewed as a good indicator for estimating external parasites increase. In addition, self-grooming time tended to be shorter in the autumn than in the summer, which seems to indicate that the cattle drove out flying insects through self-grooming or by having reduced skin temperature through moisture evaporation in the summer.
From an animal welfare point of view, it is very important to find out whether the slowdown of behavioral manifestation in the LFS was brought about from by physical discomfort, or whether the environmental adaptation is due to growth. There are many unknowns remaining about the behavioral and physiological characteristics related to the physical characteristics of beef cattle, which occur as physical growth and fattening progress simultaneously. Therefore, more advanced animal welfare for beef cattle requires further research to determine the meanings of physical and behavioral characteristics of beef cattle.
Hanwoos were found to have more changes in their behav ioral patterns in the heat of summer and within the GS group, while showing less active behavior in their LFS. These findings suggest that extra care is necessary during their the GS and LFS and during the summer period. Therefore, it is necessary to conduct animal welfare rearing management on these periods utilizing behavioral monitoring of cattle through additional behavioral physiology experiment design and more accurate behavioral analysis.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







