




INTRODUCTION
The pig (Sus scrofa domesticus) has been a major domestic animal species as an important animal protein source for humans and is also an excellent biomedical animal model related to obesity and energy metabolism in humans [1]. Korean native pigs (KNP) include those raised on the Korean peninsula as well as the Jeju native pigs raised on Jeju Island. Although Jeju native pigs, which are black in coat color, possess the disadvantages such as low feed efficiency, slow growth rate, and high backfat thickness (BFT), their meat is characterized by its solid fat tissue, white fat color, juiciness red meat color, and excellent marbling (Hereafter, the Jeju native pig is referred to as KNP). However, studies of the genetic factors that affect these meat quality characteristics have been limited in KNP.
Several genes are known to affect meat quality through different roles in meat quality characteristics: the ryanodine receptor 1 (RYR1) [2], liver fatty acid-binding protein (L-FABP) [3], obesity-associated (FTO) gene [4], melanocortin receptor type 4 (MC4R) gene [5,6], POU1F1 [7ŌĆō9], and protein kinase, AMP-activated and gamma 3 non-catalytic subunit (PRKAG3) [10ŌĆō12].
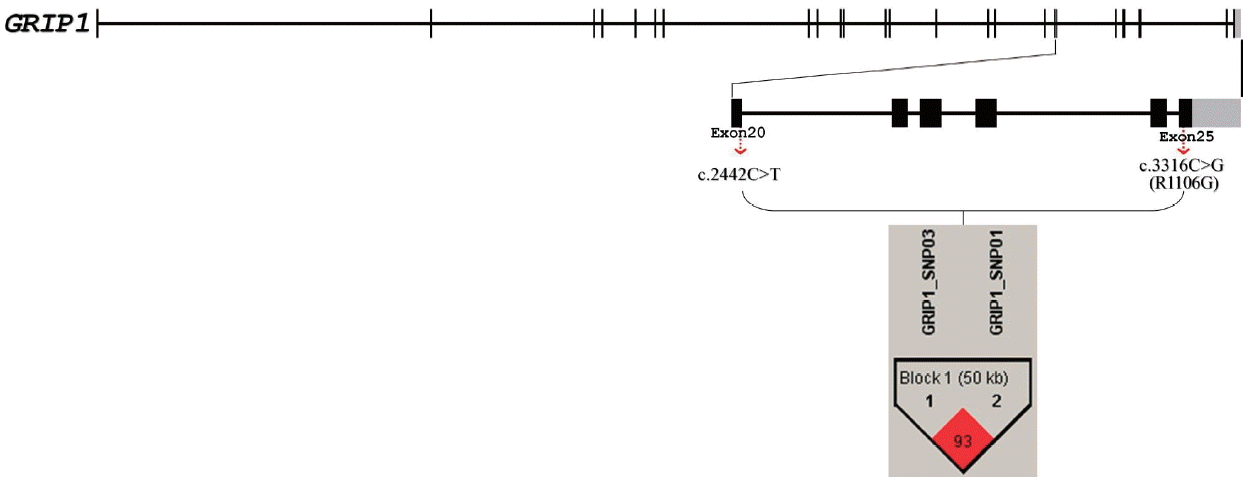
Previously, we identified a highly significant quantitative trait loci (QTL) affecting BFT traits in pig chromosome (SSC5) [13]. This QTL region (SW1482 - SW963) includes the glutamate receptor-interacting protein 1 (GRIP1) gene. Glutamate receptor interacting protein 1 is encoded by the GRIP1 gene in human [14]. GRIP1 can directly interact with and control kinesin heavy chains in dendrites as a motor for the ╬▒-amino-3-hydroxy-5-methylisoxazole-4-propionate (AMPA) receptor [15]. Glutamate receptor interacting protein 1 is encoded by the GRIP1 gene in human [14]. GRIP1 can directly interact with and control kinesin heavy chains in dendrites as a motor for the AMPA receptor [15]. GRIP1 plays a critical role in transmission in GABAergic and glutamatergic synapses [16,17]. Wu et al [18] reported that glutamatergic signaling is involved in a neuronal circuit mediating control of feeding behavior and body weight in mice [18]. Therefore, the association between single nucleotide polymorphism (SNP) markers within the exons of GRIP1 and BFT-related traits in pigs is worthy of investigation.
In this study, we presents the association of BFT traits with the GRIP1 gene using an F2 intercross population between KNP and Landrace pigs.
MATERIALS AND METHODS
Animals and measurement of BFT traits
All experimental processes, including breeding, feeding, and phenotype measurements, were conducted under identical conditions in the windowless house at the National Institute of Animal Science (NIAS, RDA) in Jeju Island. A three-generation resource population was produced and maintained as described by Cho et al [19]. Briefly, 19 purebred KNPs were crossed with 17 purebred Landrace. From these parental crosses, 91 F1 progeny and 1,105 F2 progeny (568 males and 537 females) were generated. None of the F2 males were castrated. The Animal Care Committee at the Jeju National University approved all procedures of experiments. From the F2 progeny, we measured BFT between the 4th and 5th ribs (BF_tho4_5), the 11th and 12th ribs (BF_tho11_12), and the last rib and first lumbar vertebrae (BF_tho_lum). The fat thickness was measured in millimeters.
Polymorphism detection and genotyping
Pools of genomic DNA samples from the parental lines (KNP [n = 5] and Landrace [n = 5]) of the F2 intercross were re-sequenced to detect SNP markers across the whole genome, including the GRIP1 gene (ENSSSCG00000000478), using the HiSeq2000 massively parallel sequencing technology (Illumina Inc., San Diego, CA, USA). We identified a total of 8 exonic SNP markers in the porcine GRIP1 gene. The 8 SNP markers were confirmed through pyrosequencing, using the primer sequences presented in Table 1. Among these, a non-synonymous SNP marker (c.3316 C>G [R1106G]) and synonymous SNP marker (c.2442 T>C) were found to be polymorphic in the study population. Thus, these SNP markers were genotyped in all of the F2 offspring (n = 1,105), using pyrosequencing (Biotage AB, Uppsala, Uppland, Sweden).
For the pyrosequencing analysis of missense and silent mutations in the GRIP1 gene, we made a primer set and minisequencing primer to be used in polymerase chain reaction (PCR) (Supplementary Table S1). Reactions were performed using 50 ng of genomic DNA, 0.4 ╬╝M of each primer, 1.5 mM of MgCl2, reaction buffer (Genetbio, Daejeon, Korea), 0.2 mM of dNTPs, and Taq DNA polymerase (Genetbio, Daejeon, Korea) in a 25 ╬╝L reaction volume. The PCR conditions were as follows: 35 cycles each of 30 s at 94┬░C (denaturation), 45 s at 63┬░C or 66┬░C (annealing), and 45 s at 72┬░C (elongation), 2 minutes at 94┬░C (denaturation). The genotype of the PCR products was analyzed using a pyrosequencer.
Statistical analysis of phenotype data and association test
The descriptive statistical analysis and ascertainment of normality of BFT traits were conducted using MINITAB Release 14 (MINITAB Inc., State College, PA, USA). We applied a mixed-effects linear model to perform SNP association analyses between the genetic markers and traits with simultaneous consideration of familial structure in an F2 intercross between Landrace and Korean native pigs:
Where, Yijklmn is the measured phenotype; ╬╝ is the population mean, Si is the fixed effect of ith sex, Bj is the fixed effect of jth batch, Pk is the fixed effect of kth parity, Gl is the fixed effect of lth genotype of GRIP1, b1 is a regression coefficient, CWijklm is the covariate for the carcass weight, Aijklm is the random additive polygenic animal effect and ╔øijklm is the random residual effect. The mean and variance for random additive polygenic animal effects can be defined as: A~N(0, AŽāa2), where A is based on the relationship matrix computed from the F2 intercross in this study and Žāa2 is the additive polygenic variance. The mean and variance for the residual random effect of individuals can be defined as: ╔ø~ N(0, IŽāe2), where I is the identity matrix and Žāe2 is the residual variance [20].
The significance level of the fixed effects in MODEL 1 was computed by the Wald procedure implemented in ASReml-R. The p-values of less than 0.05 were considered as statistically significant and p-values of less than 0.001 were considered as statistically highly significant. To estimate genotypic values and their standard error of each SNP genotype, the predict command in ASReml-R was used [21]. In order to estimate the additive and dominance coefficients of each SNP found in GRIP1, Gl in the Model 1 was replaced by additive variable Xadditive (coded as 1 for rare allele homozygote, ŌłÆ1 for common allele homozygote, and 0 for heterozygote), and dominance variable Xdominance (coded as 0 for two homozygotes, 1 for heterozygote). Then, trait values were regressed onto the additive and dominance coefficients at each SNP marker. The PedPhased program based on the pedigree structure was used to infer the haplotypes of GRIP1 in the F2 population [22].
RESULTS AND DISCUSSION
We previously reported QTLs affecting BFT traits on SSC5 [13]. The support interval region of the QTLs estimated by the 1.5-LOD drop method was SW1482 - SW963. In this study, a positional candidate gene analysis was performed to test whether GRIP1 in the support interval region can affect BFT traits. For the descriptive statistics, the mean┬▒standard error of the BFT traits were 34.02┬▒0.24 (BF_tho4_5), 27.99┬▒0.23 (BF_tho11_12), and 26.15┬▒ 0.22 (BF_tho_lum).
Search for mutation at the GRIP1 gene
We identified 8 putative nucleotide substitutions in GRIP1 in the exon regions using HiSeq2000 massively parallel sequencing technology (Supplementary Table S1). Among these eight putative substitutions, we detected one missense substitution [c.3316 C>G (R1106G)] and seven synonymous exonic nucleotide substitutions. To conduct the association analysis, we selected c.3316 C>G (R1106G) and c.2442 T>C because the other six substitutions were invariant, as shown in (Supplementary Table S2).
The genotyping results of c.3316 C>G (R1106G) and c.2442 T>C obtained using pyrosequencing are shown in (Supplementary Figure S1). Data on the identified genotypes and allele frequencies of c.3316 C>G (R1106G) and c.2442 T>C in GRIP1 are presented in (Table 1). The Haploview results indicated that the alleles of the two SNPs were highly associated, so it was expected that the significant results would be very similar each other (Figure 1).
Liu and Cull-Candy [23] reported that the dynamic regulation of AMPA receptors by GRIP and protein interacting with C-kinase-1 (PICK) provide a mechanism for controlling the Ca2+ permeability of the synaptic receptors. Wu et al [18] reported that glutamatergic signaling is implicated in a neuronal circuit mediating regulation of feeding behavior and body weight in mice [18]. Hence, we investigated the association between SNP markers within the exons of GRIP1 and BFT-related traits in pigs.
Association of the GRIP1 gene with BFT traits
The BFT traits were measured in more than 1,010 F2 animals. A linear mixed-effects model accounting for familial relatedness was used to evaluate the associations between each SNP marker and the BFT traits in the F2 intercross population. These two SNP markers were significantly associated with BFT between the 4th and 5th thoracic (p value = 0.00096 for c.3316 C>G [R1106G]; p value = 0.00022 for c.2442 T>C), BFT between the 11th and 12th thoracic (p value = 6.84├Ś10ŌłÆ5 for c.3316 C>G [R1106G]; p value = 9.62├Ś10ŌłÆ6 for c.2442 T>C), and BFT between the last thoracic and first lumbar vertebrae (p value = 7.92├Ś10ŌłÆ12 for c.3316 C>G [R1106G]; p value = 1.99├Ś10ŌłÆ14 for c.2442 T>C) (Table 2).
The highly significant two SNPs showed additive effects on BF_tho4_5 and BF_tho11_12, as no significant dominant effect was observed. In case of BF_tho_lum, however, the c.3316 C>G and c.2442 T>C showed both additive and dominance effects. Regarding the SNP c.3316 C>G, C3316 allele was associated with higher values of BFT values. For the SNP c.2442 T>C, T2442 allele showed higher value than C2442 allele.
Following the genome-wide linkage analysis to identify QTLs for BFT traits, a positional candidate gene analysis was conducted using an intercross between KNP and Landrace pigs. We screened the GRIP1 full-length exons for polymorphisms and detected a novel association between the missense mutation in the GRIP1 gene (c.3316 C>G [R1106G]) and BFT traits (Table 2). The change from glycine (GGA) to arginine (CGA) in exon 25 likely affects the function of GRIP1. To assess the importance of this SNP, we conducted multiple protein sequence alignment analysis [24] using human (ENSP00000381098), horse (ENSECAP00000006920), dog (ENSCAFP00000000554), cattle (XP_002687693.1), and pig (ENSSSCP00000000508, reference sequence) proteins, as well as our KNP sequences (Figure 2). The result showed that the nucleotide involved in the c.3316 C>G (R1106G) missense mutation is highly conserved among these species, providing further evidence for the putative functional importance of this SNP. However, any effect of the missense mutation (c.3316 C>G [R1106G]) on BFT traits must be validated in other independent populations prior to commercial application.
Haplo-genotype effects of the two SNPs on BFT traits are shown in Table 3. A total of seven haplo-genotypes were identified, and these haplo-genotypes exhibit different effects. The association results showed that the haplo-genotypes significantly affected BFT traits (BF_tho4_5, BF_tho11_12, and BF_tho_lum).
In this study, porcine GRIP1 was evaluated as positional candidate gene for BFT traits. We screened the GRIP1 exon regions to identify polymorphic DNA markers, and two SNPs (c.2442 T>C, c.3316 C>G [R1106G]) were found to significantly affect BFT traits in an F2 intercross population between Landrace and KNP. GRIP1 plays an important role in transmission in GABAergic and glutamatergic synapses [16,17]. After validating its functional implications in obesity related traits and this positive associations in other independent populations, the markers in GRIP1 can be used for the selection of individuals with desirable BFT traits and can contribute the understanding genetic structure of BFT traits.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print







