




INTRODUCTION
As a red carotenoid pigment without pro-vitamin A activity, astaxanthin (ASX) is a naturally occurring biological antioxidant found mainly in a wide variety of plants, algae, and seafoods (Nishigaki et al., 2010). Belonging to the carotenoid group and closely related to ╬▓-carotene, lutein, and zeaxanthin, ASX shares many of the general metabolic and physiological activities of other antioxidants. ASX has unique chemical properties of the hydroxyl (OH) and keto (C=O) moieties on each ionone ring, which explains some of its unique features, namely, the high antioxidant activity (Hussein et al., 2006).
In recent years, a number of in vitro and in vivo studies have demonstrated the antioxidant effects of ASX. Interestingly, the antioxidant activity of ASX is a result of its quenching effects on singlet molecular oxygen, strong scavenger effects on superoxide hydrogen peroxide and/or hydroxyl radicals, and inhibitory effects on lipid peroxidation (Miki, 1991; Palozza and Krinsky, 1992). In addition, studies have shown that ASX also has several other biological activities, including anticancer (Jyonouchi et al., 2000; Kurihara et al., 2002; Hix et al., 2004; Vine et al., 2005), anti-inflammatory (Ohgami et al., 2003; Choi et al., 2008), and antidiabetic effects (Uchiyama et al., 2002; Naito et al., 2004). Furthermore, ASX has beneficial effects on the skin (Lyons and OŌĆÖBrien, 2002), reproduction (Comhaire and Mahmoud, 2003), and blood pressure (Hussein et al., 2005a, b). While the effects of ASX in mammals have not been well characterized, ASX has also been shown to prevent aflatoxin-induced toxicity in mule ducklings (Cheng et al., 2001). Thus, ASX supplementation may have benefits for poultry production sustainability.
A major recurrent problem facing the poultry industry is the incidence of proventricular lesions, including proventricular enlargement and/or other pathologies associated with proventriculitis (Bayyari et al., 1995). These pathologies can result in decreased feed efficiency, decreased weight gain, and increased carcass contamination from gastrointestinal rupture during processing. Similar observations have been attributed to a number of causative agents, including biogenic amines (Barnes et al., 2001), mycotoxins (Pegram and Wyatt, 1981), copper sulphate (Wideman et al., 1995), and an unidentified filterable infectious agent (Bayyari et al., 1995), although it is likely that none of these agents is solely responsible for proventriculitis.
Biogenic amines, such as histamine (His), cadaverine, putrescine, spermine, and spermidine, are breakdown products that are produced during decarboxylation of amino acid catabolism. Biogenic amines have been shown to decrease feed efficiency (Brugh and Wilson, 1986; Stuart et al., 1986) and induce proventricular hyperplasia (Tucker et al., 1975), thereby compromising growth performance (Barnes et al., 2001). His also acts to stimulate gastric acid secretion from the parietal cells through the H2 receptors, and if dietary His exceeds the capacity for degradation, it may cause continuous stimulation of gastric acid secretion with subsequent gizzard erosion (Masumura et al., 1985). In humans, His affects circulation, gastric acid secretion, neurotransmission, and cell growth. Physical and psychological stresses also induce gastric damage, and many reports have demonstrated that reactive oxygen species cause intestinal ulceration (Das and Banerjee, 1993).
Several studies have investigated the effects of vitamins C and E against gastric ulcers caused by a variety of oxidative damages. Ascorbic acid (vitamin C) has been reported to spontaneously degrade His in the presence of molecular oxygen and copper ions (Chatterjee et al., 1975; Uchida et al., 1989). In species that are reliant on dietary vitamin C, such as humans and guinea pigs, His levels in tissues are inversely related to vitamin C status (Chatterjee et al., 1975; Clemetson, 1980; Johnston and Huang, 1991). Some studies have examined the potential interactions between ASX and natural antioxidants. The combination of ASX and ╬▒-tocopherol ameliorates oxidative injury through suppression of oxidative stress induced by diabetes (Nakano et al., 2008). Moreover, the antioxidant synergism of vitamin E and ASX was supported by decreased malondialdehyde levels (Bell et al., 2000). Thus, ASX would be expected to be a useful antioxidant for the prevention of oxidative stress. For example, ASX inhibits stress-induced impairment of antitumor activity in NK cells via its antioxidative properties, which may explain its inhibitory effects on cancer metastasis (Kurihara et al., 2002).
Despite the importance of ASX in normal physiological function and its potential involvement in pathological conditions, few studies have examined the possible interaction between His and ASX. Therefore, the objective of the current study was to investigate whether ASX had protective effects against His-induced lesions in the gizzard and proventriculus of broiler chicks. Markers of growth performance (body weight gain, feed intake, and feed conversion ratio), lesions (incidence and scores), and oxidative stress (lipid peroxidation and antioxidant enzymes) were evaluated as indicators of His exposure.
MATERIALS AND METHODS
Birds and housing
One-day-old broiler chicks (Ross 308) were obtained from a commercial hatchery (PyungTaek, Korea). They were housed in electrically heated incubators with water and a commercial broiler diet available ad libitum until they were used for experiments. At the beginning of each experiment, 48 chicks were selected from a two-fold larger population to obtain uniformity of body weight among the birds. Light was provided 23 h/d throughout the experimental period. All animal experiments were conducted in accordance with established guidelines and were approved by the Animal Care and Use Committee of Korea University.
Diets and experimental design
A commercial broiler starter diet (20% crude protein and 3,180 kcal metabolizable energy/kg) was used as a basal diet (BD). Birds were fed a BD, a diet containing 0.4% His (BD+His), a diet containing 100 ppm ASX (BD+ASX), or a diet of 100 ppm ASX plus 0.4% His (BD+ASX+His) from 5 to 26 days of age ad libitum. The ASX content of the freeze-dried Phaffia yeast was 5.0 mg/g yeast. Four 5-day-old chicks were allocated to each of the four experimental treatment groups, and the experiment was replicated three times. Doses of His and ASX were based on preliminary dose estimation.
Sampling
At the end of the experiment, the chicks and feed were weighed to determine body weight gain and feed consumption during the experiment. Blood was collected from the wing vein with heparin as the anticoagulant and centrifuged for 15 min at 1,500├Śg. The plasma was stored at ŌłÆ70┬░C for later analyses. All chicks were sacrificed by cervical dislocation, and the organs were weighed. The proventriculus and gizzard were removed and fixed in 10% buffered formalin for histology and histochemistry analyses.
Chemical analyses
ASX content in the plasma and tissue was analyzed according to the methods described by Akiba et al. (2001). Briefly, 200 ╬╝L plasma and 0.5 g tissue were added to a hexane-ethanol (1:1 v/v) solution and centrifuged at 1,500 rpm for 10 min. Cleared supernatants were collected, filtered, and injected directly for analysis by high-performance liquid chromatography (HPLC). ASX purchased from Sigma (98% purity; St. Louis, MO, USA) was used as an external standard.
Plasma vitamin E was analyzed according to the methods described by Desai (1984). In brief, 200 ╬╝L plasma was added to 2 mL of 2% pyrogallol, mixed, and incubated at ŌłÆ70┬░C for 30 min. After addition of 1 mL distilled water and 4 mL hexane, the mixture was centrifuged at 1,500 rpm for 20 min. Cleared supernatants were then dried with nitrogen gas, dissolved in hexane, and subjected to HPLC for analysis.
Tissue vitamin E was also analyzed according to the methods described by Desai (1984). Briefly, tissue was homogenized using 5 volumes of 1.15% KCl, and 1.0 mL ethanol, 0.5 mL of 25% ascorbic acid, and 1 mL of 10 N KOH were then added. The mixture was incubated at ŌłÆ70┬░C for 30 min. After adding 4 mL hexane, the mixture was centrifuged at 1,500 rpm for 10 min, and cleared supernatants were analyzed using HPLC. The mobile phase was composed of methanol and water (98:2) applied at a flow rate of 1 mL/min, and the detection wavelength was 293 nm.
Plasma and tissue vitamin C concentrations were analyzed as described previously (Omaye et al., 1979). In brief, either 400 ╬╝L plasma or 0.4 mL tissue homogenate was added into ice-cold 10% trichloroacetic acid, mixed, and centrifuged at 3,500 rpm for 20 min. Next, 500 ╬╝L of the supernatant was added into 0.1 mL 2,4-dinitrophenylhydrazine/thiourea/copper sulphate solution and incubated at 37┬░C for 3 h. After further addition of 0.75 mL of 65% sulphuric acid, the absorbance was measured at 520 nm. Plasma and liver superoxide dismutase (SOD) activities were analyzed by the formazan-WST method (Total SOD Assay Kit; Dojindo Laboratories, Tabaru, Japan). SOD activity was calculated as follows: percent inhibition (%) = ([Ablank ŌłÆ Asample]/Ablank)├Ś100 where Ablank and Asample are the absorbance values of the blank and sample, respectively.
Lipid peroxidation was determined as thiobarbituric acid reactive substances (TBARS) using the HPLC-fluorescence method. TBARS concentrations were determined from a standard curve established with TBA-malondialdehyde (MDA, 1,1,3,3-tetramethoxypropane) adducts.
For histological studies, small pieces of proventriculus and gizzard were embedded in paraffin blocks. The sections were then stained with hematoxylin-eosin dye, as described previously (Sibilia et al., 2003) followed by light microscopy observations at 10├Śmagnification. For immunohistochemistry, the proventriculus and gizzard were removed, cut into sagittal slices (4 mm thick), and fixed overnight with 10% formalin solution.
The fixed proventriculus and gizzard slices were routinely processed, embedded in paraffin, and sectioned to 7-╬╝m thickness. Hematoxylin-eosin staining was performed to examine histopathological changes. Sections were deparaffinized in xylene, rehydrated in a graded alcohol series, incubated in a solution of 3% hydrogen peroxide in methanol for 30 min, and microwaved at 750 W for 10 min in 10 nM citrate buffer (pH 6.0). Nonspecific binding sites were blocked with normal rabbit serum diluted 1:10 in phosphate-buffered saline (PBS) for 30 min and incubated overnight at 4┬░C with primary antiserum. After rinsing in PBS, sections were incubated in biotinylated secondary antibody (Vector Laboratories, Burlingame, CA, USA) for 30 min. The antigen-antibody complex was visualized by an avidin-biotin-peroxidase complex solution (Vector Laboratories, USA) with 3,3-diamino benzidine (Invitrogen, Carlsbad, CA, USA). Tissue sections were then rinsed in distilled water and counter stained with MayerŌĆÖs hematoxylin.
RESULTS
Body weight gain, feed intake, and feed conversion ratios are summarized in Table 1. Birds fed a diet supplemented with ASX had the highest body weight gain and feed intake, while those consuming His ate less and weighted the least. However, birds consuming a diet containing both His and ASX exhibited the least growth efficiency in terms of body weight gain and feed intake. Addition of ASX to the diet caused chicks to consume less feed per gram gain than other treatments. These data indicated that birds consuming ASX were able to convert feed to growth more efficiently.
With regard to organ/weight ratio, none of the diets affected the fresh weights of the liver, spleen, heart, and fabricius (Table 2). Supplementation with His resulted in enlargement of the proventriculus, gizzard, pancreas, and abdominal fat, and the addition of ASX did not alleviate His-induced organ enlargement.
The plasma concentration of vitamin C was not affected by supplementation with ASX or His, whereas hepatic vitamin C level increased more with ASX than other supplementations (Table 3). Dietary ASX or His had lowering effects on plasma and liver vitamin E concentrations compared to the dietary BD.
The effects of dietary supplementation with ASX or His on lesions associated with proventriculus are shown in Table 4. Higher lesion scores were consistently observed in chicks consuming dietary His compared with those consuming the BD. Interestingly, supplementation with ASX resulted in a lower incidence of lesions and reduced lesion scores. When the BD was supplemented with ASX and His, however, the effects of ASX on reducing the incidence of proventricular lesions was not obvious.
SOD and TBARS are extensively used as biochemical indicators of the pathological states associated with oxidative stress and lipid peroxidation, respectively. His supplementation caused a reduction in SOD levels in the blood, while the opposite effect was observed for dietary ASX (Table 5). In contrast, none of the dietary treatments caused significant alterations in SOD activities in the liver. His supplementation significantly altered lipid peroxidation in the liver, while ASX did not prevent the increase in MDA (Table 5). Neither His nor ASX changed the level of MDA in the blood.
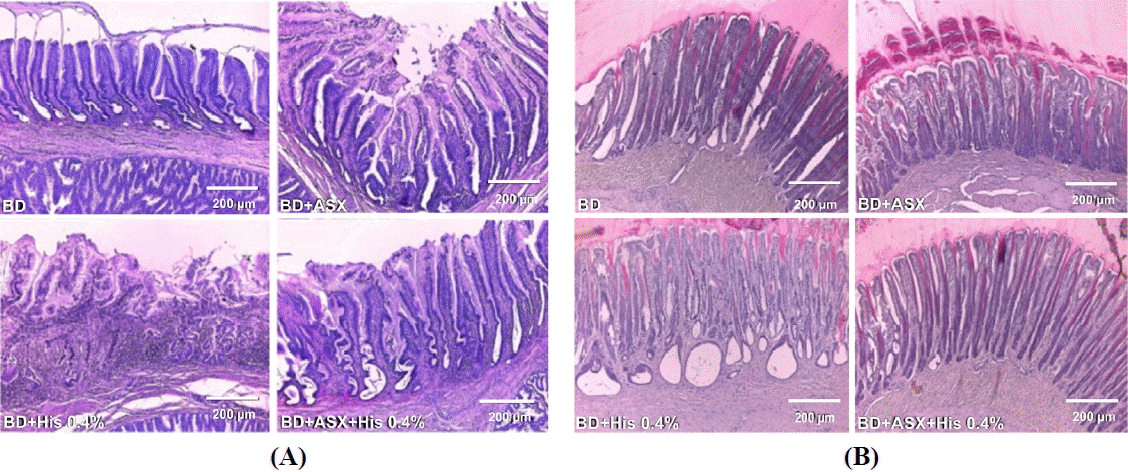
Figure 1A shows the extent of lesions in the mucosal layer of the proventriculus stained with hematoxylin-eosin. While no mucosal damage was observed in the BD group, His caused severe mucosal ulcerative lesions in the mucosal epithelium and lamina propria of the proventriculus. In contrast, in chicks treated with ASX, unique degeneration and necrosis of the epithelium were not observed. Further examination of the proventriculus with SOD immunoperoxidase staining (figure not shown) revealed that ASX reduced the His-dependent increase in SOD expression in the gastric mucosal layer. The extent of lesions in the mucosal layer of the gizzard is shown in Figure 1B. In contrast to the results in the BD group, His supplementation resulted in the formation of severe lesions of vesicular enlargement in the mucosa of the gizzard. ASX treatment led to recovery of normal vesicles.
DISCUSSION
In this study, we investigated whether ASX had protective effects against His-induced lesions in the gizzard and proventriculus of broiler chicks. Our data demonstrated that ASX lowered the frequency of proventricular ulceration induced by His, indicating that ASX may be useful for alleviating structural damage to the digestive system in poultry under certain stressful conditions.
ASX, one of the most common carotenoids, exerts powerful antioxidant properties by cleavage of singlet oxygen. Therefore, ASX protects many tissues from lipid peroxidation in mammals (Goto et al., 2001; Hussein et al., 2006). However, our current results indicated that feeding a diet supplemented with ASX and His was not effective for the prevention of lipid peroxidation estimated by MDA production, as measured by determining TBARS levels. On the other hand, antioxidative enzymes, such as SOD, can scavenge radicals and contribute to the bodyŌĆÖs defenses against oxidative stress. In particular, SODs catalyze the dismutation of superoxide and hydrogen peroxide, the first step of enzymatic antioxidative defenses. Dietary supplementation with ASX and His resulted in a significant inhibition of plasma SOD activity compared with that of His supplementation alone. Hence, increased SOD expression may neutralize the harmful effects of superoxides from His during the early stress response in the plasma. Contrary to the findings that exogenously administered ASX can be incorporated into microsomes of the liver tissue and other organs in male broiler chicks (Takahashi et al., 2004), ASX was not incorporated into the liver in our study, and thus, did not prevent lipid peroxidation in chicks, probably due to the gender factor of chicks and the preparation of ASX.
Growth performance was significantly increased by supplementation with ASX in the current study. Although data describing the effects of ASX on growth performance are conflicting, it is possible that ASX stimulates feed intake via its known anti-inflammatory effects. In fact, Cheng et al. (2001) found that ASX alleviated growth retardation by enhancing the chemotaxis ability of macrophages under aflatoxin-induced toxicity in mule ducklings. This theory was further supported by the enhancement of antibody production and proliferation of T cells as the centrepiece of immune function by ASX feeding (Takimoto et al., 2007). However, our data describing the growth performance of chicks showed that supplementation with ASX was not efficacious in the feed conversion ratio when the diet was supplemented with His. While the reason for this discrepancy is not clear, ASX supplementation may be associated with species-specific immune and anti-inflammatory functions.
His is known to increase gastric acid secretion and causes enlargement of the proventriculus (Harry et al., 1975). Consistent with previous reports, we found that His consumption resulted in increased proventricular weight and lesions with concurrent reductions in body weight and increased feed conversion. For example, Harry et al. (1975) and Barnes et al. (2001) fed chicks His at 0.1% to 1.0% and 0.1% to 0.2%, respectively, and observed decreases in body weight and feed intake and an increase in the incidence of proventricular enlargement. Our data demonstrated that His induced several pathologies associated with proventriculitis of the glandular epithelium, fibrosis, or oedema. Interestingly, enlargement of the proventriculus was further related to inhibition of abdominal fat accumulation. ASX was found in high amounts in the liver, confirming that the liver was the main organ of metabolic activity, where ASX promotes expression of cytochrome P450 to facilitate hepatic drug metabolism (Takimoto et al., 2007). Moreover, His prevented ASX from being absorbed, resulting in reduced levels of ASX in the liver. Since our study involved a relatively small sample, our data may still be open for interpretation, particularly the data for SOD and TBARS. As with all studies involving ASX, the responses of various types of animals to the treatment may differ.
In conclusion, dietary supplementation with ASX influenced growth performance but did not affect lipid peroxidation in broiler chickens. Furthermore, ASX might be helpful to alleviate structural damages to the digestive systems of poultry under certain stressful conditions.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







