




INTRODUCTION
The hypothalamus plays a pivotal role in integrating external environmental cues and generates the appropriate responses to influence feed intake in mammals, as well as in birds (Schwartz et al., 2000). As in mammals, a population of neurons that influence appetite exist in the hypothalamus of avian species: Orexigenic neurons (e.g., neuropeptide Y [NPY]) (Schwartz et al., 2000) and anorexigenic neurons (e.g., proopiomelanocortin [POMC], corticotropin-releasing factor) (Richards, 2003; Boswell, 2005). Peripheral signal reflecting energy balance can be integrated at the hypothalamic arcuate nucleus in mammals and infundibular nucleus in avian species.
Insulin has multiple functions in insulin receptor (INSR)-expressing tissues (hypothalamus, fat, muscle, liver) (Saltiel and Kahn, 2001; Shiraishi, et al., 2011a). The INSRs are also widely distributed in the central nervous system, and regulate vital physiological processes in mammals (Pagotto, 2009). Intracerebroventricularly (ICV) injection of insulin can decrease feed consumption in layer chicks when compared with control (Shiraishi et al., 2011b). Honda et al. (2007) demonstrated that central insulin was as an appetite-suppressive peptide in chicken central nervous system, and this anorexigenic action was mediated by POMC and corticotropin-releasing hormone (CRH). But the regulation of peripheral insulin on appetite in chickens remains unknown. In the present study, we examined the changes in feed intake and appetite-related peptides in the hypothalamus of broiler chicks after subcutaneous insulin injection and determined the relationship between peripheral insulin and appetite.
MATERIALS AND METHODS
Animals
Male broiler (Arbor Acres) chicks were obtained from a local hatchery at 1 day of age and reared in an environmentally controlled room. Temperature and lighting was maintained according to commercial conditions. All chicks received a starter diet containing 21.5% crude protein and 12.33 MJ/kg of metabolizable energy. All birds had free access to feed and water. This study was approved by the Shandong Agricultural University and carried out in accordance with the “Guidelines for Experimental Animals” of the Ministry of Science and Technology (Beijing, China).
Experimental protocol and sample collection
Experiment 1. Effects of insulin on feed intake in chicks under feeding condition
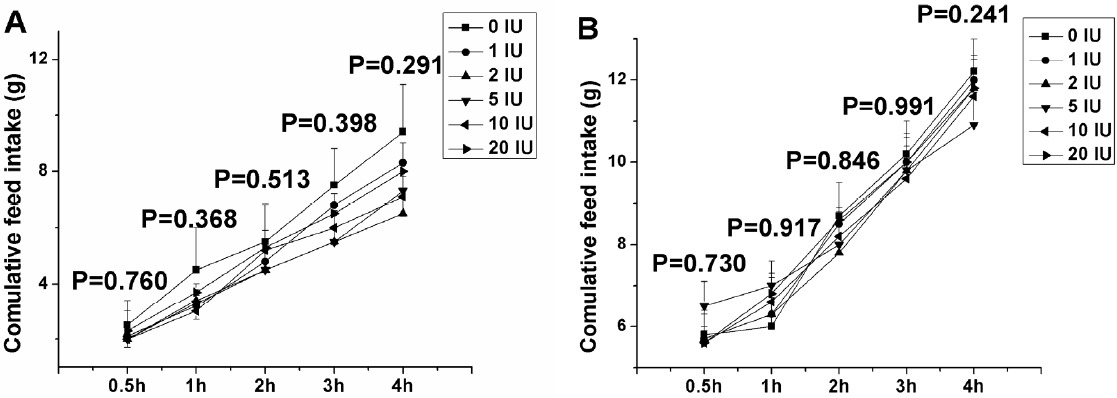
A total of 72, 6-d-old chicks of similar body weight were divided into 6 groups, 6 replicates per group, and 4 chicks per replicate. The chicks with ad libitum feeding received a subcutaneous injection of 0, 1, 3, 5, 10, or 20 IU of insulin or vehicle (saline). Food intake was recorded in 0.5, 1, 2, 3, and 4 h after injection. A blood sample was drawn from a wing vein using a heparinised syringe for 30 s and collected in iced tubes 4 h after injection. Plasma was obtained following centrifugation at 400 g for 10 min at 4°C and stored at −20°C for subsequent analysis of glucose and insulin.
Experiment 2. Effects of insulin on feed intake in chicks under fasting condition
A total of 72, 6-d-old chicks of similar body weight were divided into 6 groups, 6 replicates per group, and 2 chicks per replicate. The chicks, after fasting for 3 h, received a subcutaneous injection of 0, 1, 3, 5, 10, or 20 IU of insulin or vehicle (saline). Feed intake was recorded 0.5, 1, 2, 3, and 4 h after injection. A blood sample was collected 4 h after injection. The treatment of control and 10 IU insulin were repeated twice. During the second experiment, the chicks were continuously fasted for 2 or 4 h after peripheral insulin administration, and the hypothalamus samples were collected according to Yuan et al. (2009). After being flash-frozen in liquid nitrogen, the samples were stored at −80°C for subsequent RNA extraction.
Measurement of plasma glucose and insulin levels
Plasma glucose concentration was measured spectrophotometrically with commercial diagnostic kits (Hitachi High-Technologies Corp.; Jiancheng Bioengineering Institute, Nanjing, China) that have been successfully applied in poultry research (Wang et al., 2012).
Plasma insulin was measured by radioimmunoassay with a guinea pig anti-porcine insulin serum (3V Bio-engineering group Co., Weifang, China). In this procedure, 125I-labelled porcine insulin competes with chicken insulin for sites on anti-porcine insulin antibodies that are immobilised on the wall of a polypropylene tube. Significant cross-reactivity has been observed between chicken insulin and guinea pig anti-porcine sera (Simon et al., 1974). The insulin in this study is referred to as immunoreactive insulin. The sensitivity of the assay was 1 μIU/mL, and all samples were included in the same assay to avoid interassay variability. The intraassay coefficient of variation was 2.88%.
RNA isolation and analysis
Total RNA extraction and quantitative real-time polymerase chain reaction (PCR) were performed as described previously (Liu et al., 2014). Sequences of primers are shown in Table 1. The PCR data were analyzed with the 2−ΔΔCT method (Livak and Schmittgen, 2001). The mRNA levels of target genes were normalized to 18S and β-actin (ΔCT). On the basis of the Ct values, 18S and glyceraldehydes-3-phosphate dehydrogenase (GAPDH) mRNA expression were stable across the treatments in this study (p>0.1).
Statistical analysis
The data are presented as means±standard error of the mean. Homogeneity of variances among the treatments was confirmed using Bartlett’s test. All data were subjected to one-way analysis of variance analysis to test the main effect of the treatment (n = 8). When the main effect of the treatment was significant, the differences between means were assessed by Duncan’s multiple range analysis. Less than 0.05 p value was considered statistically significant.
RESULTS
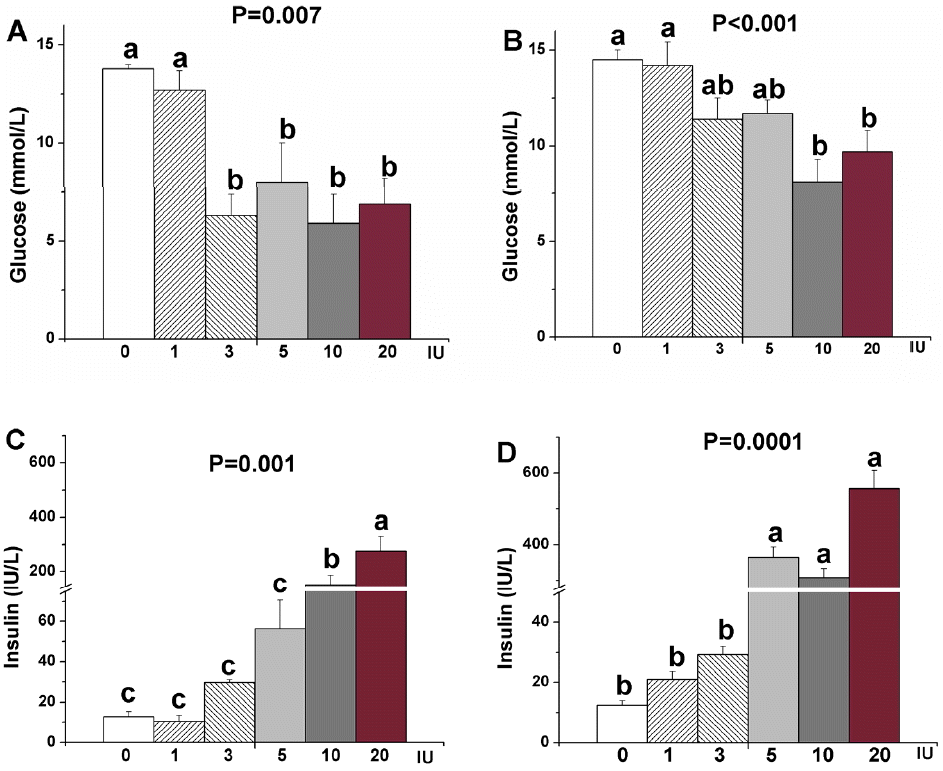
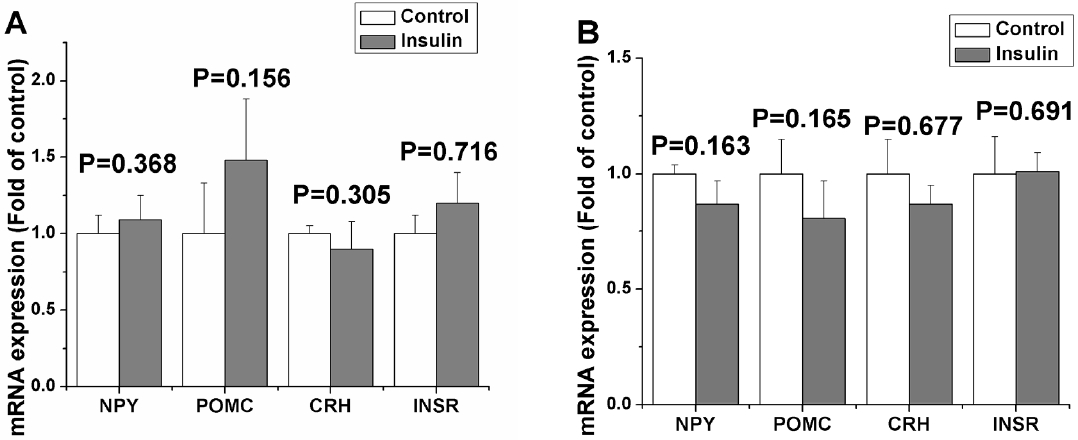
Peripheral insulin treatment (1 to 20 IU) did not significantly alter the feed intake of chicks either under ad libitum feeding or fasting conditions in 0.5, 1, 2, 3, and 4 h (p>0.05). Compared with the control, plasma glucose concentration was significantly decreased 4 h after insulin treatment of 3, 5, 10, and 20 IU in chicks under ad libitum feeding (p<0.05). But the increased plasma insulin concentration was only observed in the doses of 10 and 20 IU in chicks under ad libitum feeding (p<0.05). In fasted chicks, 4 h after 10 and 20 IU insulin treatment the plasma glucose concentration was significantly decreased (p<0.05), but not following the dose of 1, 3, and 5 IU (p>0.05). Peripheral insulin treatment (1 to 20 IU) induced a dose-dependent increase in insulin concentration of plasma in fasted chicks (p<0.05). Two or 4 h after peripheral insulin treatment of 10 IU there was no significant effects on the hypothalamic genes expression of NPY, POMC, CRH, and INSR.
DISCUSSION
In the present study, we investigated the effect of peripheral insulin treatment on appetite of chicks. Our data demonstrated that i) Peripheral insulin injection dose not significantly affect the feed intake of chicks; ii) Peripheral insulin treatment dose not significantly alter the hypothalamic INSR and appetite-related genes expression of NPY, POMC, and CRH. The results suggest that peripheral insulin injection does not alter appetite of chicks.
In mammals, changes in the circulating level of insulin signal the hypothalamus to effect long-term changes in energy balance by activating and/or inhibiting specific anabolic and catabolic pathways (Schwartz et al., 2000; Woods et al., 2006). Obici et al. (2002) found that insulin could decrease appetite via INSRs in mammals. Although INSRs have been identified in the chicken brain (Simon and Leroith, 1986), the effect of peripheral administration of insulin on appetite in birds is not clear. Because plasma insulin levels rise and fall with increases and decreases in energy state in chickens such as during fasting and refeeding, it is possible that peripheral insulin could serve as peripheral signal of energy state in birds as they have been postulated to do in mammals (Niswender et al., 2004; Woods et al., 2006). But the feed intake was not changed by peripheral insulin treatment in fasted and fed chicks, which is consistent with the previous observation of ICV injection of insulin-induced no change in the feed consumption in boiler chicks under fasting and ad libitum conditions (Shiraishi et al., 2011b). But the result is inconsistent with the study in layer chicks and sheep. Thirty min following an ICV insulin injection the feed consumption in layer chicks was significantly inhibited under both fasting and ad libitum conditions (Shiraishi et al., 2011b). And 1 h after the intrajugular administration of insulin the feed intake of sheep was significantly depressed (Deetz and Wangsness, 1980). These conflicting results suggest that insulin resistance exists of broiler chicks, possibly due to persistent hyperinsulinemia, which results in a down-regulation of central nervous system INSR expression (Shiraishi et al., 2011b).
Chickens have a blood glucose level that is twice as high as that of most mammals, and can survive massive doses of insulin that would be lethal in mammals (Gibson et al., 1980). So chickens are regarded as an insulin resistant animal. In our experiment, the high dose insulin changed plasma glucose level, but not low doses. Therefore, large doses of insulin are required for stimulation of glucose uptake in insulin responsive tissues in chickens (Gomez-Capilla and Langslow, 1977). The regulation of blood glucose in chickens is unorthodox and not well understood, yet glucose is an important signal of energy or nutritional status that is integrated into the regulation of feed intake and energy metabolism in mammals. Transient change in plasma glucose level caused by insulin did not alter feed intake in chicks, which is consistent with the previous study in chickens (Simon et al., 2000), but not in mammals (Branton et al., 2014). Therefore, the role of insulin, as an afferent signal for energy stores in broiler chickens remains to be determined.
Blood insulin is thought to cross the blood-brain barrier to act within the brain to help control appetite via blinding INSR. In neonatal chicks, immunostaining revealed hypothalamic neuron expressing INSRs in the infundibular nucleus (the avian equivalent of the mammalian arcuate nucleus), paraventricular nucleus, ventromedial hypothalamus and lateral hypothalamus (Shiraishi et al., 2011a). However, INSR levels were not affected by feeding condition in broiler chicks, but were affected in layer chicks (Shiraishi et al., 2011b). In our experiment, peripheral insulin did not significantly affect hypothalamic INSR levels. The results imply that hypothalamic INSR are not sensitive to the insulin in fat-line chickens, and it may be an explanation why insulin resistance or typrII diabetes easily occurs in obese animals.
NPY has been shown to be a potent orexigenic agent in chickens when administered centrally (Kuenzel and McMurtry, 1988). ICV NPY injection can elevate the plasma insulin level in broiler chicks (Kuenzel and McMurtry, 1988). In addition, hypothalamus-damaged cockerels appeared hypertrophy of pancreatic B cells, hyperactivity of insulin secretion and hyperphagia (Sonoda, 1983). But we did not detect any change of hypothalamic NPY mRNA levels, which is inconsistent with ICV insulin injection in chicks (Shiraishi et al., 2011a) and with ICV insulin injection in rats (Schwartz et al., 1992). The results suggest regulation of insulin on hypothalamic NPY is subject to species diversity. In addition, the effect of central NPY on peripheral insulin is non-reversible in chicks.
Different from NPY, hypothalamic POMC neurons play an important role in transducing the afferent input, i.e., hormonal and metabolic signals, into a neuronal anorexigenic response in both chickens and mammals. Hypothalamic POMC mRNA levels are increased by peripheral insulin and decreased by fasting in mammals (Benoit et al., 2002). Although central insulin administration could up-regulate POMC mRNA level in chicks (Honda et al., 2007), fasting for 48 h (Song et al., 2012) or peripheral leptin (Dridi et al., 2005), insulin administration has no significant effect on hypothalamic POMC mRNA levels (Song et al., 2012). The results suggest that hypothalamic POMC of chick is insensitive to peripheral metabolic signals.
ICV CRH injection significantly decreased feed intake in both feeding and overnight-fasting broiler chickens and laying hens (Denbow et al., 1999). Chan et al. (2005) has reported that peripheral insulin administration increases CRH mRNA level in rat hypothalamus. Hypothalamic CRH mRNA level in chicks hypothalamus is significantly increased by central insulin administration (Honda et al., 2007), but not by peripheral insulin injection. All results imply that the effect of insulin is not only associated with species, but also with treatment method (e.g., central and peripheral administration).
CONCLUSION
Peripheral insulin (1 to 20 IU) administration didn’t alter the feed intake in chicks under ad libitum and fasting conditions. And hypothalamic INSR, POMC, CRH, and NPY genes were also not changed by peripheral insulin administration. All results suggest peripheral insulin administration has no effect on appetite in chicks.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







