




INTRODUCTION
Intestinal epithelium not only allows selective permeability as a filter for absorption of nutrients, but also is regarded as the first line of defense against foreign antigens, such as pathogens and toxins coming from intestinal lumen (Niessen and Gottardi, 2008). Tight junction (TJ) is an important mode of cell-to-cell adhesions in the epithelial and endothelial cellular sheets, which acts as a primary barrier that prevents the diffusion of solutes through the intercellular space, especially in the intestine (Gonz├Īlez-Mariscal et al., 2008). In animals, including pigs (Smith et al., 2010; Zhao et al., 2011), the TJ and barrier function of intestine is crucial at the time of weaning, since the TJ can be damaged easily by bacterial toxins (Wijtten et al., 2011; Hu et al., 2013). Lipopolysaccharide (LPS) is one of the important bacterial toxin which can induce stress (Li et al., 2012; Guo et al., 2013). Therefore, improvement of TJ formation is considered as one of the most important steps to form an intestinal mucosal barrier against paracellular diffusion of microorganisms, toxins, and other harmful antigens across the epithelium. Glucagon-like peptide-2 (GLP-2) is an intestinotrophic growth hormone which can enhance the biological function of the intestinal. Researches showed that GLP-2 can significantly improve the villus height and weight of the small intestine in piglets and improve digestion and absorption enzyme expression in piglet intestinal (Naimi et al., 2013; Sigalet et al., 2014; Thymann et al., 2014). In addition, GLP-2 could recover and maintain the integrity of the intestinal mucosa epithelium barrier by increasing the expressions of zonula occludens protein 1 (ZO-1), claudin-1 and occludin in obstructive jaundice rats (Chen et al., 2008). GLP-2 could modulate the intestinal cellsŌĆÖ proliferation and apoptosis via the phosphatidylinositol 3-kinase (PI3K)-protein kinase B(Akt)-mammalian target of rapamycin (mTOR) signaling pathway (Cheeseman, 1997; Yusta et al., 2002). However, there are no studies of GLP-2 regulating the expression of TJ proteins in the IPEC-J2 cells. Therefore, the present experiments were conducted to test the hypothesis that GLP-2 could promote TJŌĆÖs expression and transepithelial electrical resistance (TER) in LPS stressed and normal IPEC-J2 cells and GLP-2 could regulate TJ and TER through the PI3K-Akt-mTOR signaling pathway in IPEC-J2 cells. This research can help to understand the molecular mechanism of intestinal damage and intestinal barrier function changes and can also provide the basis for the future application of GLP-2 to improve the animal intestinal barrier function.
MATERIALS AND METHODS
The Chinese Council on Laboratory Animal Care protocols and guidelines were followed when caring for the animals.
Reagents
DulbeccoŌĆÖs modified Eagle medium (DMEM:HamŌĆÖs F-12 [1:1]), fetal bovine serum, Penicillin-Streptomycin Antibiotic Mixture and insulin-transferrin-selenium-X (ITS-X) were from GIBCO BRL (Gibco BRL, Gaithersburg, MD, USA). GLP-2 was from Phoenix pharmaceuticals, Belmont, CA, USA. LPS, wortmannin, LY294002, and all other chemical reagents were from Sigma Chemical (Sigma Chemical Co. St. Louis, MO, USA). Antibodies (anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH), occludin, claudin-1, ZO-1, phosphorylated Akt [p-Akt], and phosphorylated mTOR [p-mTOR]) were from Santa Cruz.
Cell culture
Under sterile conditions, non-transformed IPEC-J2 cell line (College of Life Science, University of Copenhagen in Denmark) was cultured in DulbeccoŌĆÖs modified Eagle medium (DMEM:HamŌĆÖs F-12 [1:1]) supplemented with 5% fetal bovine serum, 1% insulin-transferrin-selenium-X (ITS-X) and antibiotics in an incubator with atmosphere of 5% CO2 at 37┬░C. The medium was replaced every 24 h. To examine the effect of GLP-2 in the process of occludin, claudin-1, and ZO-1 mRNA expressions, 5├Ś105 cells were cultured on 6-well plate for 3 days until reaching the confluence. For the electrical resistance experiment, the cells were grown on 0.4-╬╝m polyester membrane trans-well (PET, 4.67 cm2, 0.4 ╬╝m, Coning, 3450) at a density of 5├Ś104 cells/well for 3 days. To test the effect of GLP-2 in the process of occludin, claudin-1 and ZO-1 proteins expressions, 1├Ś106 cells were cultured on 100 mm culture dish for 4 days. The test time is 24 hours. The results of optimal concentration test showed that the optimal concentration of GLP-2 and LPS were 100 nmol/L and 100 ╬╝g/mL. The four treatments were control group, 100 nmol/L GLP-2 -treated group, 100 ╬╝g/mL LPS-treated group and 100 ╬╝g/mL LPS with 100 nmol/L GLP-2-treated group. Once the improvement effects of GLP-2 on the TJ proteins and TER in IPEC-J2 was established, wortmannin and LY294002 were used to test the molecular mechanism. The other four treatments were control group, 100 nmol/L GLP-2-treated group, 100 nmol/L GLP-2 with 10 nmol/L wortmannin and 100 nmol/L GLP-2 with 10 ╬╝mol/L LY294002-treated group. Kinase (Akt, mTOR) assay cultivating time was 45 minutes and the others were 24 hours. The method of cell culture with the front line.
Transepithelial electrical resistance measurement
IPEC-J2 cells TER across the monolayer was measured by placing one of the electrodes in the upper well and the other in the lower well using a Millicell Electrical Resistance System with EVOM2 (15148-AB01D, World Precision Instruments, Sarasota, FL, USA) according to the manufacturerŌĆÖs instructions. Two readings were taken per well and averaged. The resistance from each well was subtracted by its appropriate blank, the inserts without cells to arrive at the resistance of the monolayer and then multiplied by the effective growing area to obtain TER (Ohm*4.67 cm2).
mRNA expressions measurement
After being washed by pre-cooling PBS 3 times, TRIZOL reagent (TaKaRa, Dalian, Liaoning, China) was added to samples of IPEC-J2. RNA extraction was performed using the chloroform/isopropyl alcohol method, followed by Qiagen RNeasy clean-up, according to the manufacturerŌĆÖs instructions. RNA was eluted from the column in 30 ╬╝L of RNase-free water, and concentration and purity of RNA were determined using a DU-800 spectrophotometer (Beckman, Kraemer Boulevard, Brea, CA, USA). Reverse transcription polymerase chain reaction (PCR) was performed using the PrimeScript RT reagent Kit with gDNA Eraser (Perfect Real Time) (Code No. RR047A; TaKaRa, China). Reaction conditions and PCR program followed the manufacturerŌĆÖs instructions. Different porcine gene specific primers (Invitrogen, Carlsbad, CA, USA) were used (Table 1). Primer pairs were designed using Primer3 program based on NCBI porcine sequences. Real time PCR (RT-PCR) was performed using a CFX-96 real time PCR system (Bio-Rad, Alfred Nobel Drive, Hercules, CA, USA) with the SYBR Premix Ex Taq TM II (Tli RNaseH Plus) (Code No. RR820A; TaKaRa, China). cDNA (1 ╬╝L) was added in a 25 ╬╝L reaction mixture which contained 12.5 ╬╝L SYBR Premix Ex Taq II (Tli RNaseH Plus) (2├Ś), 10 ╬╝M each primer, 2 ╬╝L cDNA and 8.5 ╬╝L dH2O. The program setting was 30 s at 95┬░C and then 40 cycles of 95┬░C for 5 s, 60┬░C to 62┬░C (depending on the primers used) for 30 s, and 72┬░C for 30 s. On the completion of amplification, a melting curve was produced by holding the reaction mixture at 65┬░C to 95┬░C with a heating rate of 0.1 ┬░C per second and a continuous fluorescence measurement and finally a cooling step to 40┬░C. The housekeeping gene encoding GAPDH was used as an internal control. The analysis consisted of four independent experiments. Each experimental sample was assayed using four replicates for each primer. The relative levels of the mRNA of the genes of interest were normalized to the GAPDH mRNA and were determined using a modified 2ŌłÆ╬ö╬öCt method (Pfaffl, 2001).
Protein preparation and western blotting
IPEC-J2 cells were washed with ice-cold PBS and immediately lysed in lysis buffer (20 mM Tris-HCl, 150 mM NaCl, 1 mM Ethylenediaminetetraacetic acid (EDTA), 1% Triton X-100) followed by quantitation of protein using Micro BCA kit (Thermo, Rockford, IL, USA). After the protein concentration of supernatant fractions was quantized by a standard bicinchoninic acid (BCA) protein assay kit (Bio-Rad, USA), equal amount of proteins for each sample was loaded in 10% Tris-glycine polyacrylamide gels and electrophoresed. Then, the proteins were transferred onto a polyvinylidene difluoride microporous membrane (Millpore, Bedford, MA, USA) for 2 h at 4┬░C and blocked with 5% skim milk in TBS-T (20 mM Tris HCl, 100 mM NaCl, 0.05% Tween 20) for 90 min and then incubated sequentially with primary antibody overnight. The primary antibodies being used were: Polyclonal anti-claudin-1 (Santa Cruz Biotechnology Inc., Dallas, TX, USA) diluted 1:500, anti-occludin (Santa Cruz Biotechnology Inc., USA) diluted 1:450, anti-ZO-1 (Santa Cruz Biotechnology Inc., USA) diluted 1:400, anti-p-Akt (Ser473) (Santa Cruz Biotechnology Inc., USA) diluted 1:1,000, anti-pmTOR (Ser2448) (Santa Cruz Biotechnology Inc., USA) diluted 1:2,000 and anti-GAPDH (Cell Signaling Technology Inc., Danvers, MA, USA) diluted 1:2,000. Subsequently, the membrane was washed and incubated with IgG-HRP (Santa Cruz Biotechnology Inc., USA ) (diluted at 1:5,000) for 1 h. The target proteins were visualized by enhanced chemiluminescence system (GE Healthcare, Waukesha, WI, USA) followed by analysis using Gel Doc TM XR (Bio-rad, USA). Intensity of the blotting was quantified using quality-one software.
Statistical analysis
Sigmastat statistical software (SPSS17.0) was used to analyze densitometry results of TER, mRNA and protein relative expression levels for Western blots. All data were reported as means┬▒standard deviation from 3 independent experiments. One-way analysis of variance (ANOVA) was used to determine whether a significant difference was present among all treatment groups. Additionally, Bonferroni t tests were performed for comparisons when the ANOVA was very significant at p<0.01 (Agresti and Kateri, 2011).
RESULTS
Effects on transepithelial electrical resistance
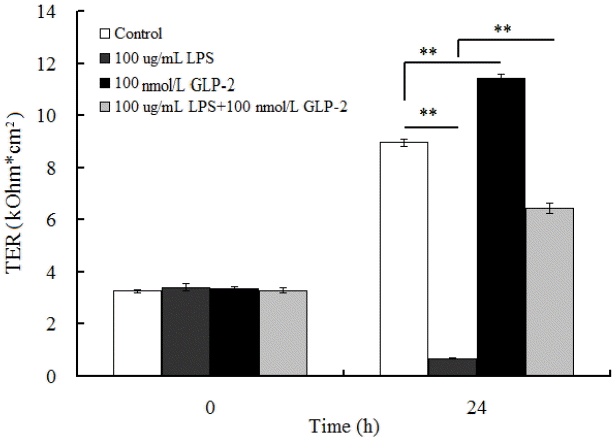
Compared with the control group, 100 ╬╝g/mL LPS significantly decreased the IPEC-J2 cellŌĆÖs TER 92.4% (p<0.01). IPEC-J2 cellŌĆÖs TER significantly increased by 27.2% in 100 nmol/L GLP-2-treated group (p<0.01). GLP-2 inhibited the damnification of TER caused by LPS stress and maintained the barrier function of IPEC-J2 cells (Figure 1).
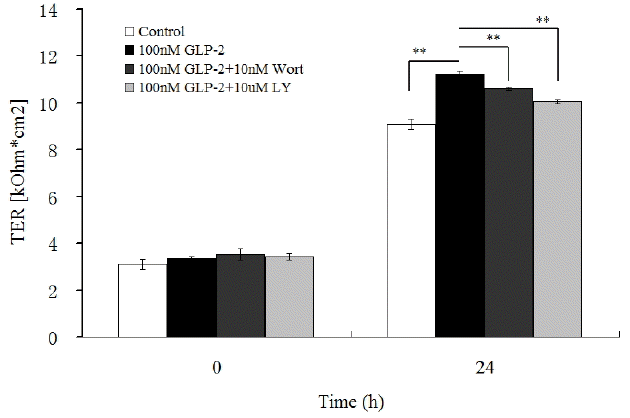
As shown in Figure 2, in comparison with the control group, GLP-2 (100 nmol/L) increased the TER (p<0.01). But when wortmannin (10 nmol/L) was added into the medium, the TER significantly decreased by 5.4% (p<0.01) and decreased by 10.3% in the 100 nmol/L GLP-2 with 10 ╬╝mol/L LY294002-treated group (p<0.01).
Effects on mRNA expressions of tight junction and signaling molecules
The results showed that compared with the control, the expressions of occludin, claudin-1 and ZO-1 mRNA decreased by 46%, 57%, and 34% respectively in the 100 ╬╝g/mL LPS -treated group (p<0.01) and increased by 148%, 261%, and 54% respectively in the 100 nmol/L GLP-2-treated group (p<0.01). Compared with the 100 ╬╝g/mL LPS-treated group, 100 ╬╝g/mL LPS with 100 nmol/L GLP-2 significantly increased occludin, claudin-1, and ZO-1 mRNA expression (p<0.01) respectively by 46.3%, 65.1%, and 30.3% (Table 2).
As depicted in Table 3, compared with the control, the addition of GLP-2 (100 nmol/L) increased the expressions of Akt, mTOR, occluding, claudin, and ZO-1 mRNA (p<0.01) while these effects were blocked when wortmannin (10 nmol/L) and LY294002 (10 ╬╝mol/L) was simultaneously added into the medium with GLP-2. Wortmannin and LY294002 inhibited GLP-2-induced up-regulation of occluding, claudin and ZO-1 mRNA expressions (p<0.05). Compared with the 100 nmol/L GLP-2-treated group, the mRNA expressions of Akt, mTOR, occluding, claudin and ZO-1 decreased by 46.9%, 50.5%, 38.1%, 49.6%, and 18.9% respectively in the 100 nmol/L GLP-2 with 10 nmol/L wortmannin-treated group (p<0.01) and decreased 67.2%, 70.8%, 49.6%, 60.9%, and 25.8% respectively in the 100 nmol/L GLP-2 with 10 ╬╝mol/L LY294002-treated group (p<0.01).
Effects on proteins expressions of tight junction and signaling molecules
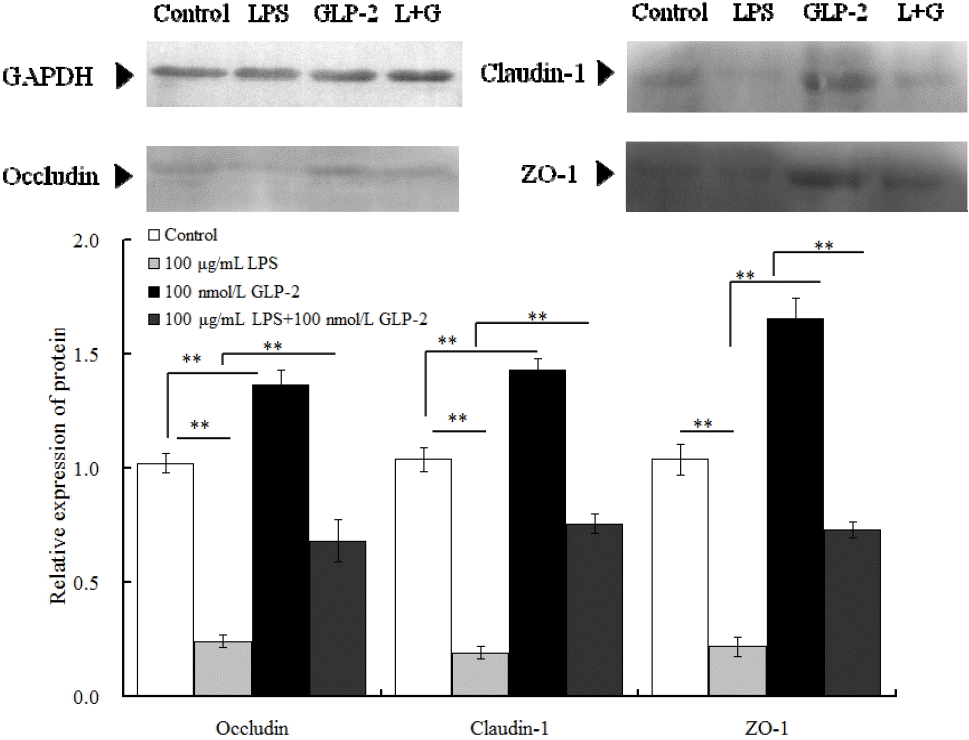
In comparison with the control group, the protein relative abundance of occluding, claudin and ZO-1 increased by 35%, 39%, and 62% respectively (p<0.01) in the 100 nmol/L GLP-2-treated group and decreased by 76.5%, 81.7%, and 78.8% respectively (p<0.01) in the 100 ╬╝g/mL LPS-treated group. However, compared with the LPS-treated group, 100 ╬╝g/mL LPS with 100 nmol/L GLP-2 significantly increased occludin, claudin-1 and ZO-1 protein expression (p<0.01) respectively by 183%, 300%, and 231.8% (Figure 3).
As shown in Figure 4, in comparison with the control group, GLP-2 (100 nmol/L) increased the abundance of all protein (p<0.01). But when wortmannin (10 nmol/L) was added into the medium, the relative abundance of p-Akt, p-mTOR, occluding, claudin, and ZO-1 significantly decreased by 15.5%, 15.4%, 10.8%, 14.2%, and 27.6% respectively (p<0.01) and decreased by 24.2%, 22.5%, 13.7%, 16.3%, and 31.9% respectively in the 100 nmol/L GLP-2 with 10 ╬╝mol/L LY294002-treated group (p<0.01).
DISCUSSION
GLP-2 is a 33-amino acid intesti-notrophic hormone derived from posttranslational processing of proglucagon in the enteroendocrine L cells of the ileum and colon (Cameron and Perdue, 2005). As a physiological regulator of mucosal epithelial homeostasis, GLP-2 plays an important role in intestinal permeability. Dong et al. (2013) found that GLP-2 can significantly improve the expression of occludin and Claudin-3, 7 mRNA and proteins in the mouse jejunum. Moran et al. (2012) found that GLP-2 can not only significantly increase the expression of occludin and ZO-1 proteins in Caco-2 cell but also can effectively inhibit the reduction of occludin and ZO-1 expression caused by tumor necrosis factor ╬▒ (TNF-╬▒) stress. The above results show that, GLP-2 can effectively promote the expression of TJ in intestinal epithelial cells and enhance intestinal barrier function. The study on piglet jejunum cell model results show that GLP-2 can significantly improve the expressions of occludin, Claudin-1 and ZO-1 in normal IPEC-J2 cells. In addition, GLP-2 can also inhibit TJ expression decreasing caused by LPS stress in IPEC-J2 cells. The results suggest that, GLP-2 can promote healthy intestinal epithelial TJ mRNA and protein expressions and can effectively inhibit of the immune stress caused by LPS. GLP-2 may be used as a promising alleviate nutrition relieves stress of weaned piglets.
The results of study on LPS effecting on IPEC-J2 cells show that LPS can increase cell inflammation factor and induce cells inflammatory reaction (Razzuoli et al., 2013; Paszti-Gere et al., 2014). Our study found when IPEC-J2 cells were stressed by LPS, the expression level of TJ proteins occludin, claudin-1, and ZO-1 mRNA and proteins significantly decreased, which indicated that LPS could cause serious damage to IPEC-J2 cellsŌĆÖ TJ. Hou et al. (2012) study found that LPS could significantly reduce expressions of claudin-1 and occludin in weaned pigletsŌĆÖ jejunum and ileum epithelial cells and can reduce the content of RNA and total protein in jejunum. In addition, Liu et al. (2008) found that, LPS could change intestinal morphology and increase intestinal epithelial permeability and destroy the intestinal barrier function. We also found that the expression of 100 ╬╝g/mL LPS could not completely destroy IPEC-J2 cell TJ protein and the cell barrier function. The results were in accord with the experimental results reported by Geens and Niewold (2010), and Arce et al. (2010), indicating that IPEC-J2 cells have a certain resistance to LPS immune stress and can protect the cell structure and function from being completely damaged by LPS immune stress. The early research showed that GLP-2 can increase the content of sodium dependent glucose transporter protein in mouse jejunal brush border membrane and can promote the growth and development of jejunum. The further results showed that GLP-2 promote cell growth through PI3K when PI3K specific inhibitor Wortmannin was used (Cheeseman et al., 1997). Yusta et al. (2002) found that GLP-2 can promote baby hamster kidney epithelial cell growth though the PI3K-Akt signal pathway when use LY294002 as PI3K specific inhibitor. Shi et al. (2011) found that GLP-2 can promote protein synthesize in epithelial cells through PI3K-Akt-mTOR signal pathway and this result determine the important role of mTOR in the process that GLP-2 promote protein expression in epithelial cells. However, there were articles reporting that GLP-2 didnŌĆÖt modulate the mouse or human colon cancer cell growth and survival through the PI3K-Akt-mTOR signal transduction pathway (Koehler et al., 2008). The study suggests that GLP-2 regulate the growth, intracellular protein expression through the cell regulation of PI3K-Akt-mTOR signal transduction pathway is not in all cells. Therefore, it is significant to illustrate the role of PI3K-Akt-mTOR signal transduction pathway in the GLP-2 regulation of TJ in piglets jejunum epithelial cells. This experiment results showed that GLP-2 can significantly increase the expression of occludin, claudin-1 protein, and ZO-1 through PI3K-Akt-mTOR signal transduction pathway. The results was in accord with Shi et al. (2013), suggesting that PI3K-Akt-mTOR signal transduction pathway is one of important signaling pathways that GLP-2 regulate cell TJ and barrier function.
CONCLUSION
In summary, the results showed that LPS stress significantly reduced expression of TJ in IPEC-J2 cells and damage the barrier function of IPEC-J2 cells. GLP-2 can increase TER on the LPS stressed IPEC-J2 cells and enhance the barrier function, which are associated with the functions of promoting the mRNA and proteins expressions of the key proteins of TJ (occludin, claudin-1, ZO-1). GLP-2 may modulate TJŌĆÖs expression and barrier function through PI3K-Akt-mTOR signal transduction pathway in IPEC-J2 cells.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







