




INTRODUCTION
Reactive oxygen species (ROS) can induce embryo damage through oxidative stress, including two-cell block by lipid peroxidation, enzyme inactivation, nuclear DNA fragmentation, mitochondrial damage, adenosine triphosphate (ATP) depletion, and embryo apoptosis. ROS are generated during oxidative phosphorylation, and are removed by non-enzymatic (e.g., antioxidant vitamins) and enzymatic defense systems (e.g., superoxide dismutase) (reviewed in [Guerin et al., 2001]). They occur more frequently in high-oxygen environments (Goto et al., 1993), which typically have insufficient defense mechanisms to prevent ROS-induced damage (Kitagawa et al., 2004).
The reduction of ROS is important for preventing oxidative damage to embryos in vitro. To this end, studies have suggested that using co-culture systems to reduce oxygen concentrations from 20% to 5% during in vitro embryo development (Kitagawa et al., 2004), and decreasing light exposure (Goto et al., 1993) can protect embryos against oxidative stress. In addition, various methods of treatment, including the addition of antioxidants (e.g., resveratrol, a natural antioxidant) to porcine embryo cultures, have also shown promise in reducing oxidative stress (Kitagawa et al., 2004; Lee et al., 2010a). Ginsenoside is a major pharmacological component of ginseng (Lu et al., 2009), and can be divided into two groups: Rb1 and Rg1. Ginsenoside Rg1, from Panax ginseng C. A. Meyer, has various pharmacological actions and multiple effects in vivo, including anti-diabetic (Xie et al., 2005), anti-amnestic (Mook-Jung et al., 2001), and immunity-boosting effects (Kenarova et al., 1990), as well as antioxidant (Chen et al., 2003) and proliferative activities (Shi et al., 2009). However, its effects on embryo development have not been investigated.
In this study, we investigated the effects of ginsenoside Rg1 on in vitro porcine embryo development. To this end, we used in vitro-produced parthenogenetic-activated porcine embryos, and analyzed the effects of ginsenoside Rg1 on the total cell number and rates of cleavage, formation, and apoptosis of blastocysts. The mechanisms underlying these effects were also examined. Surprisingly, ginsenoside Rg1 improved porcine embryo quality, not through antioxidant function, but by stimulating metabolic pathways, particularly the glucose uptake pathway.
MATERIALS AND METHODS
Animal welfare
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional guides on the care and use of laboratory animals. The care and experimental use of pigs was approved by the Institute of Laboratory Animal Resources, Seoul National University (SNU-140328-2).
Chemicals
Ginsenoside Rg1 (purity ≥98% by high performance liquid chromatography) was purchased from Ambo Institute (Daejeon, Korea), and stored at −20°C until use. It was mixed with porcine zygote medium 3 (PZM-3), and diluted before use. All of the chemicals were purchased from Sigma-Aldrich Corp. (St. Louis, MO, USA) unless otherwise stated.
Oocyte collection and in vitro maturation
Porcine ovaries, donated by the Sooam Biotech Research Institute (Seoul, Korea), were used. Porcine follicular fluid (PFF) and cumulus-oocyte complexes (COCs) were aspirated using a syringe and 18-gauge needle. Sediments were washed twice with Tyrodes lactate-Hepes medium supplemented with 0.01% polyvinyl alcohol (TL-Hepes-PVA). Oocytes with compact cumulus cells and granulated cytoplasm were selected for in vitro maturation (IVM). Selected COCs were washed twice with TL-Hepes-PVA medium and cultured in IVM medium (TCM-199; Life Technologies, Rockville, MD, USA) supplemented with 10 ng/mL epidermal growth factor, 1 μg/mL insulin, and 10% (v/v) PFF for 44 h at 39°C in 5% CO2 and humidified conditions. COCs were treated with hormones, 4 IU/mL equine chorionic gonadotropin (eCG; Intervet, Cambridge, UK), and human chorionic gonadotropin (hCG; Intervet, UK) for the first 22 h, and then were maturated in hormone-free conditions for the last 22 h. After 44 h of maturation, oocytes were isolated from cumulus cells using 0.1% (w/v) hyaluronidase. Cumulus-free oocytes were washed twice with Dulbecco’s phosphate-buffered saline (DPBS; Welgene, Daegu, Korea) supplemented with 0.4% (w/v) bovine serum albumin (BSA).
Parthenogenetic activation
Cumulus-free oocytes were placed in an electrode chamber, and electronically activated by an electric pulse (1.0 kV/cm for 60 μs) in activation solution (280 mM mannitol, 0.01 mM CaCl2, 0.05 mM MgCl2) using a BTX Electro cell manipulator ECM2001 (BTX, San Diego, CA, USA). Parthenogenetic-activated oocytes were washed twice with 0.4% (w/v) BSA-supplemented DPBS (Welgene, Korea), followed by a 4 h incubation in PZM-3-containing 2 mmol/L 6-dimethylaminopurin for post-activation.
In vitro culture and ginsenoside Rg1 treatment
Post-activated oocytes were cultured in drops of PZM-3 covered with mineral oil for 168 h. Embryo culture conditions were maintained at 39°C in a humidified atmosphere, 5% CO2, and 5% O2 for the positive control group, and 20% O2 for the treatment and negative control groups. Ginsenoside Rg1 was used at concentrations of 5, 10, 20, and 50 μg/mL. The cleavage rate was calculated between 48 to 72 h. The number of blastocysts and the rate of blastocyst formation were calculated 168 h after parthenogenetic activation.
Cell counts and apoptosis measurements
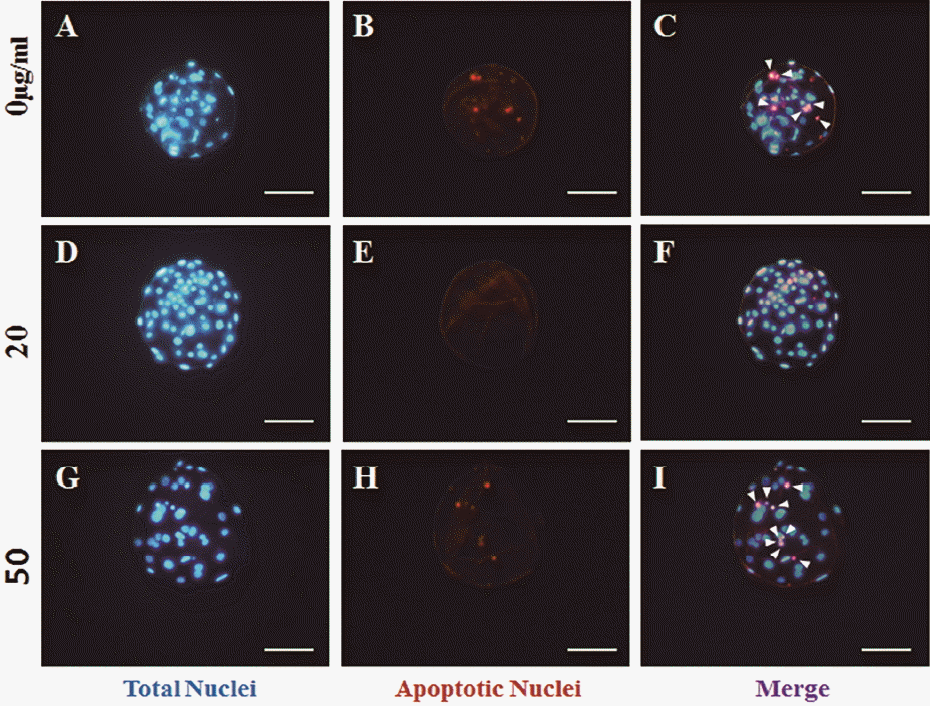
Blastocysts cultured for 168 h were fixed in 4% (w/v) paraformaldehyde for 1 h. After fixation, an in situ cell death detection kit (Roche, Mannheim, Germany) was used in a terminal deoxynucleotidyltransferase (TdT)-mediated deoxy-uridine nick-end labeling (TUNEL) assay according to the manufacturer’s instructions. Then, blastocysts were stained with 5 mg/mL bisbenzimide (Hoechst 33342) for 5 min. Total cell numbers and apoptosis rates for blastocysts were calculated after staining under a UV light using a micromanipulator (eclipse TE2000-U; Nikon, Tokyo, Japan).
Analysis of mRNA expression by quantitative real-time polymerase chain reaction
The zona pellucida of unhatched blastocysts was removed with acidified Tyrode’s solution before mRNA extraction. mRNA was extracted from embryos using a Dynabeads mRNA Direct Kit (Life Technologies, USA), according to the manufacturer’s instructions. cDNA synthesis was performed using a High Capacity RNA-to-cDNA (cDNA Reverse Transcription) Kit (Applied Biosystems; ABI, Foster City, CA, USA), according to the manufacturer’s instructions. Briefly, at a final volume of 20 μL, cDNA synthesis was performed at 37.5°C for 60 min, and samples were subsequently incubated at 95°C for 5 min to activate the reverse transcription reaction. Synthesized cDNA samples were stored at −80°C until subsequent use. Quantitative real-time polymerase chain reaction (qPCR) was performed using an ABI 7300 Real-Time PCR System (Applied Biosystems, USA). A DyNAmo HS SYBR Green qPCR Kit (Thermo scientific, Rockford, IL, USA) was used for real-time quantification of the PCR products. For amplification, 0.1 μM of each primer listed in Table 1 and 0.5 μL blastocyst cDNA were added to the 10 μL reaction mixture. The reactions were performed under the following conditions: 1 cycle at 95°C for 10 min; 40 cycles at 95°C for 15 s; and 60 s at annealing temperature. The dissociation curve was analyzed to confirm the specificity of the PCR product. Beta-actin was used as a control gene to determine the relative quantity. All of the genes were measured in triplicate, and relative expression ratios were analyzed by the 2−ΔΔCt (threshold cycle) method (Livak and Schmittgen, 2001).
Statistical analysis
All of the data were analyzed using the GraphPad Prism statistical program (GraphPad Software, San Diego, CA, USA). Cleavage rate, blastocyst formation rate, total cell number, and apoptosis rate were analyzed using one-way analysis of variance (ANOVA) and Dunnett’s test. qPCR data were analyzed using ANOVA and Fisher’s least significant difference. All of the data are reported as means±standard errors of the mean. p values less than 0.05 were considered statistically significant.
RESULTS
Effects of O2 concentration and ginsenoside Rg1 on parthenote cleavage and blastocyst formation rates
Porcine parthenotes were divided into six groups including two control groups grown under 5% or 20% O2, and four treatment groups grown under 20% O2. The cleavage rate of each group was determined 48 to 72 h after parthenogenetic activation. The embryo cleavage rate did not differ between parthenotes cultured in 5% O2 (positive control group) and 20% O2 (negative control group). Moreover, ginsenoside Rg1 had no significant effect on the embryo cleavage rate (Table 2). The rate of blastocyst development was analyzed 168 h after parthenogenetic activation, and there were no significant differences between the positive and negative control groups. Furthermore, treatment with ginsenoside Rg1 did not significantly alter the blastocyst formation rate (Table 2). These results demonstrate that O2 concentration and ginsenoside Rg1 treatment do not affect cleavage or blastocyst formation rates in porcine parthenotes cultured in vitro.
Effects of O2 concentration and ginsenoside Rg1 treatment on total blastocyst cell number and rate of apoptosis
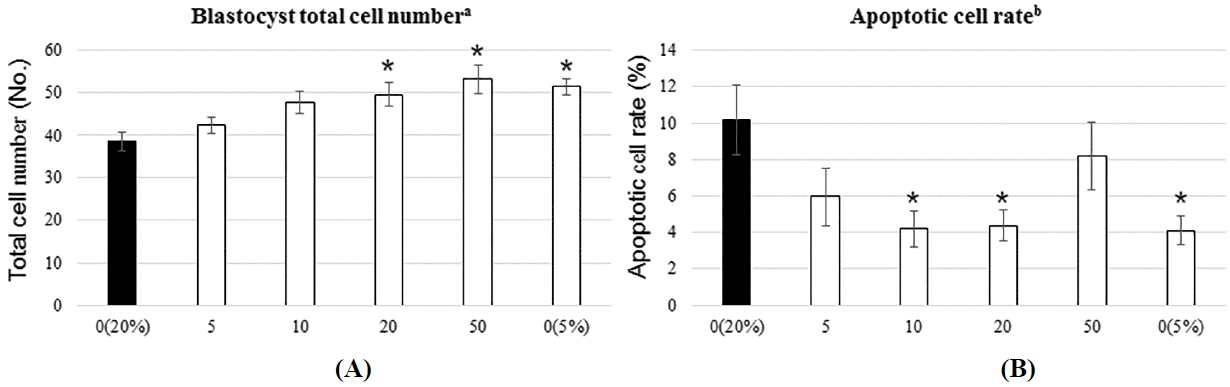
The total number of blastocyst nuclei was determined by Hoechst 33342 staining 168 h after parthenogenetic activation. Apoptotic cell rate was subsequently calculated using a TUNEL assay. The total number of blastocyst cells was significantly higher in the positive control group than the negative control group. Treatment with ginsenoside Rg1 (20 and 50 μg/mL) significantly increased cell number compared to the negative control group (Figures 1 and 2A). The rate of blastocyst cell apoptosis was significantly lower in the positive control group than the negative control group. In addition, ginsenoside Rg1 treatment (10 and 20 μg/mL) significantly decreased DNA fragmentation compared to the negative control group (Figures 1 and 2B). Treatment with 5 μg/mL ginsenoside had no effect on cell number or apoptotic cell rate, and treatment with 10 μg/mL had no effect on total cell number, but decreased the rate of apoptosis. In contrast, treatment with 50 μg/mL increased the total cell number, but had no effect on apoptosis (Figure 2). Therefore, high concentrations of ginsenoside Rg1 (50 μg/mL) do not reduce apoptosis, whereas medium concentrations (10, 20 μg/mL) do. In addition, high concentrations of ginsenoside (20 and 50 μg/mL) increase cell number, whereas lower concentrations do not. These results suggest that ginsenoside may be toxic at high concentration (50 μg/mL) (Liu et al., 2006). Together, these data demonstrate that 20 μg/mL is the best treatment concentration because it increased total cell number and decreased the rate of apoptosis.
Changes in mRNA expression levels
Due to the results of the above experiments, 20 μg/mL ginsenoside was selected for subsequent experiments. Genes related to intrinsic apoptosis pathways were selected to investigate the antioxidative effects of ginsenoside Rg1. Treatment with ginsenoside Rg1 did not change the expression of apoptosis-related genes including Bcl-2 homologous antagonist/killer (BAK1), B-cell lymphoma-extra large (Bcl-xL, BCL2-like 1 isoform 1), Caspase 3 (CASPASE3), and tumor protein p53 (TP53) compared to the negative control group. In addition, there were no differences in the expression of BAK1, Bcl-xL, TP53, and CASPASE3 between the positive and negative control groups (Figure 3A). Additional genes involved in metabolic processes were also examined. The expressions of Carnitine palmitoyltransferase 1B (CPT1B) and mechanistic target of rapamycin (mTOR), metabolism-related genes, were similar between the groups; however, ginsenoside treatment increased the expression of glucose transporter 1 (GLUT1), a glucose transporter (Figure 3B).
DISCUSSION
We investigated the effects of ginsenoside g1 treatment on in vitro-produced preimplantation embryos grown in high-oxygen conditions, which are of lower quality than in vivo-produced embryos (Mateusen et al., 2005). There are many differences between in vivo and in vitro environments, including differences in ROS. To control for these differences, various experiments designed to protect embryos from oxidative stress have been performed, including co-culture systems (Menezo et al., 1990), decreasing light exposure (Goto et al., 1993), and antioxidant treatments (Kitagawa et al., 2004; Lee et al., 2010a). Ginsenoside Rg1 is a natural antioxidant that activates internal antioxidant enzymes and scavenges free radicals (Deng and Zhang, 1991). It also stimulates glucose uptake through two pathways (Shang et al., 2008; Lee et al., 2010b). However, its effects on porcine embryo development have not yet been examined.
Apoptosis involves various pathways, including extrinsic, intrinsic, and perforin/granzyme pathways. Of these, the intrinsic pathway is induced by hazardous factors such as radiation, toxins, hypoxia, hyperthermia, viral infections, and free radicals (Elmore, 2007). BAK1, Bcl-xL, CASPASE3, and TP53 are known regulators of apoptosis, and are related to the intrinsic mitochondrial apoptosis pathway. The Bcl-2 family controls apoptotic mitochondrial events (Cory and Adams, 2002); BAK1 and Bcl-xL have roles in pro- and anti-apoptotic signaling, respectively (Elmore, 2007); Caspase3 acts during the apoptosis execution phase by initiating DNA fragmentation (Stennicke and Salvesen, 1998); and TP53 is a cell cycle regulator that can activate apoptosis (Fridman and Lowe, 2003). Despite their importance in apoptosis, their expressions were not altered by ginsenoside treatment, and did not differ between the positive and negative control groups. These results indicate that the antioxidant effect of ginsenoside on embryos is not related to the intrinsic apoptosis pathway (Figure 3A). Major metabolic pathways consisting of various specific pathways utilize energy substrates including glucose, amino acids, and fatty acids to produce ATP, and these pathways are important for early embryo development (Lane and Gardner, 1996). During embryo development, GLUT1 is the major transporter responsible for glucose uptake in the preimplantation embryo (Morita et al., 1992), and glucose is the predominant energy source during the blastocyst stage (Brison and Leese, 1991). mTOR participates in the initiation of protein synthesis, growth, and the first cell division in early development and trophoblast behavior (Land et al., 2014). CPT1B is involved in oxygen uptake as well as beta-oxidation and degradation of fatty acids (Gentile et al., 2004). The results from this study show that, compared to positive and negative controls, 20 μg/mL ginsenoside increased GLUT1 expression. Because high levels of GLUT1 correlate with increased glucose uptake (Kumar et al., 2015), these data indicate that glucose uptake was increased in ginsenoside-treated embryos. Ginsenoside stimulates glucose uptake through two pathways (Shang et al., 2008; Lee et al., 2010b), including the insulin signaling pathway by promoting the activation of both GLUT1 and GLUT4 (Shang et al., 2008). Previous studies have also shown that in mouse preimplantation embryos, glucose uptake can affect total cell number, but not blastocyst formation rates (Martin and Leese, 1995). The results of the present study correspond well with previous studies performed in mice. Two other studies have reported that in human and porcine embryo, glucose uptake is minimal during early embryo developmental stage and is increased rapidly after 8-cell stage (Conaghan et al., 1993). Glucose also did not affect the rate of blastocyst formation (Hagen et al., 1991). Thereby, ginsenoside Rg1 treatment, which increases glucose uptake did not affect early embryo development (e.g., embryo cleavage rate and blastocyst formation rate).
Taken together, our data indicate that treatment with 20 μg/mL ginsenoside Rg1 can increase the total number of blastocyst cells and decrease the apoptotic cell rate in embryos grown in high-oxygen conditions. Although the TUNEL assay showed changes in the apoptotic rate, no differences in apoptosis-related gene expression were found between the treated and untreated groups. However, ginsenoside treatment did increase GLUT1 mRNA expression, indicating increased glucose uptake, which may function to counteract damage from ROS. Thus, ginsenoside Rg1 can improve embryo quality by increasing glucose uptake in high-oxygen conditions. Previous studies have focused on decreasing ROS by treatment with various antioxidants to improve embryo development; however, this study shows that increasing glucose uptake can also improve embryo quality by protecting the cells from oxidative stress in high-oxygen conditions. Therefore, these results suggest that stimulating metabolic processes may be a potential method for reducing oxidative stress during in vitro culture of embryos.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







