



INTRODUCTION
Superovulation is a term used to describe the administration of exogenous gonadotrophin to induce the growth and maturation of many ovarian follicles to increase the number of mature ovum for fertilization (Baldassarre et al., 2003). Embryo recovery from donor and transfer of recovered embryo to synchronized recipient is considered as an effective means of increasing the contribution of superior females to the gene pool. However, the result of any superovulation programme is inconsistent and can fluctuate from complete failure to total success without any change in the standard operating procedure (Baldassarre and Karatzas, 2004). This variation in ovarian response occurs not only between treatments but also within treatment. This may be attributed by many intrinsic and extrinsic factors including season, breed, donor age, nutrition, purity and source of gonadotropin and treatment protocols (Gonzalez-Bulnes et al., 2004).
Superovulation can be performed using exogenous gonadotrophin such as equine chorionic gonadotrophin (Goel and Agrawal, 2005), horse anterior pituitary (Moore, 1974) and follicle stimulating hormone (FSH) (Abdullah et al., 2012). Due to a short half-life and better embryo production performance compared to other gonadotrophins, FSH is preferable as a stimulatory agent in any multiple ovulation and embryo transfer (MOET) programme (Holtz, 2005). However, various factors (e.g., ratio of FSH and luteinizing hormone (LH) in hormone preparations, route of administration, dosage and frequency of injection) are involved on FSH superovulatory performance (Nowshari et al., 1995; Lehloenya and Greyling, 2009; Yuan et al., 2011).
Superovulatory treatment of porcine follicle stimulating hormone (pFSH) in goat mainly consists of 200 mg pFSH without consideration of body weight (BW) of the goat, which varies from 28 to 68 kg, and also produces variable results (Menchaca et al., 2007; Lehloenya and Greyling, 2010; Xiao et al., 2013). Thus, the dosage of gonadotrophin might be a cause of variation in superovulation response. Theoretically, it seems that the goat with larger BW needs more pFSH than the smaller goat. Some researchers used different dosages of pFSH for superovulation without considering the BW of goat (Chang et al., 2006; Yuan et al., 2011; Abdullah et al., 2012) and cattle (Ali et al., 2012). Furthermore, the mature BW of goats varies among breeds. However, there is limited information regarding the use of pFSH dosage based on BW of goats. Hence, it is necessary to know the suitable dosage range according to BW for successful embryo transfer programme. Therefore, the objective of the present study was to evaluate the effect of different dosages of pFSH on ovarian stimulation and embryo production performance of Boer crossbred goats.
MATERIALS AND METHODS
Location of experiment
This experiment was carried out at the Institute of Biological Sciences Mini Farm (at a location of 2° 30′ N, 112° 30′ E), the University of Malaya, Malaysia. This location is 60 m above sea level and has an annual rainfall of 2600 mm. All animals used in this experiment were in accordance with the guidelines of the University of Malaya.
Experimental animal management
Thirty mature crossbred does (local×Boer) of 17 to 36 kg BW and 24 to 60 months of age were used for this experiment. The experimental does were divided into 3 groups and termed as pFSH-3, pFSH-5 and pFSH-8. The pFSH-3, pFSH-5 and pFSH-8 groups were administered 3, 5 and 8 mg pFSH per kg BW, respectively. The average BW of the does of pFSH-3, pFSH-5, and pFSH-8 groups were 29.50, 28.81, and 25.00 kg respectively. The experimental does were reared under an intensive management system and received fresh soya waste (20% dry matter) at a rate of 1 kg/head/d. The soya waste contained 27.9% crude protein, 30.5% neutral detergent fibre and 5.3% ash. Soya waste was offered to the animal once in the morning, while the Napier grass (Pennisetum purpureum) was offered in the morning and afternoon. All animals had free access to water and salt lick. The study was conducted between October 2012 and April 2013.
Ultrasonography of experimental animals
All the donor does were scanned to identify the pregnancy status using an ultrasound machine (ALOKA SSD 500, Hitachi Aloka Medical, Ltd. Tokyo, Japan) equipped with a 7.5 MHz transrectal probe. The tip of the probe was lubricated with carboxyethylcellulose contact gel and was gently inserted until the urinary bladder was identifiable. The probe was moved gently forward and backward and rotated at 90° clockwise and 180° counterclockwise to identify both ovaries. Only non-pregnant does were used for this experiment.
Estrus synchronization and superovulation treatments
For estrus synchronization, a controlled internal drug release (CIDR) dispenser was inserted (D0; 0800 to 0900 h) for 14 days (D14; 0800 to 0900 h) and a single injection of 125 μg prostaglandin F2α (PGF2α) (Estrumate; Intervet International B.V. Boxmeer, The Netherlands) was administered intramuscularly 1 day before starting the pFSH treatment (D11). The pFSH treatments were initiated at 2 days before the CIDR removal (D12), and were administered twice a day through multiple intramuscular injections. Subsequently, all the donor does were divided randomly into 3 groups, namely pFSH-3, pFSH-5, and pFSh-8. pFSH-3, pFSH-5, and pFSH-8 were administered with 3, 5, and 8 mg of pFSH (Folltropin-V; Bioniche Life Sciences Inc, Ontario, Canada) based on per kg BW through multiple intramuscular injections, respectively. Total amount of pFSH for each donor was divided into six decreasing (25, 25, 15, 15, 10, and 10%) dosages and given twice daily (0800 to 0900 h and 1800 to 1900 h) for a period of three successive days. Based on these percentages, the absolute amount of pFSH per kg BW was 0.75, 0.75, 0.45, 0.45, 0.30, and 0.30 mg for pFSH-3; 1.25, 1.25, 0.75, 0.75, 0.50, and 0.50 mg for pFSH-5; and 2.00, 2.00, 1.20, 1.20, 0.80, and 0.80 mg for pFSH-8, respectively. Correspondingly, the ranges of pFSH administered per doe were 60 to 123, 110 to 180, and 136 to 276 mg, for pFSH-3, pFSH-5 and pFSH-8 groups, respectively.
Estrus detection and natural mating
After CIDR withdrawal, estrus was observed 3 times in a day (morning, afternoon and evening) until exhibition of overt estrus by placing a buck of proven libido. After showing the signs of estrus, the doe was placed in a pen with a male for natural mating and AI was done simultaneously at 48 and 72 hours after CIDR removal.
Surgical procedure for ovarian responses assessment
On Day 7 after CIDR removal, ovarian responses were evaluated and embryos were collected during a laparotomy session. The donor goats were off-feed and off-water for 16 to 20 hours before surgery. For goat anesthesia, xylazine hydrochloride was mixed with ketamine hydrochloride at a ratio of 1:50 and administered through intramuscular injection (11 mg/kg BW). After anesthesia, the reproductive tract was exteriorised through a mid-ventral incision and the superovulatory responses were assessed by counting the corpus luteum (CL) and anovulatory follicles of both ovaries. Both the fallopian tubes were flushed using a flushing medium consisted of phosphate buffered saline supplemented with streptomycin, penicillin and polyvinylpyrrolidone. A two-way Foley catheter (Imex) was used for embryo flushing. The recovered structures (embryos plus unfertilized oocytes) in the flushing medium were evaluated and classified accordingly under a stereomicroscope (SZH10; Olympus Optical Co. Ltd, Tokyo, Japan).
Statistical analysis
The effect of dosages of gonadotrophin on ovarian stimulation and embryo production parameters was examined by one-way analysis of variance using SPSS software (version 16.0, Statistical Package for the Social Sciences software, Chicago, IL, USA). Regression procedure was carried out using MS Excel software to determine the relationship between body weight and estrus response. Results were expressed as the mean±standard error of mean, and differences and correlations were considered as significant when p<0.05.
RESULTS
Ovarian stimulation of crossbred does as affected by different pFSH dosages is presented in Table 1. Donor does showed estrus after 24 hours upon CIDR removal. All the donor does (100%) of pFSH-5 and pFSH-8 showed signs of standing estrus, however, half of the donor does of pFSH-8 showed signs of standing estrus. Average numbers of CL for pFSH-3, pFSH-5, and pFSH-8 were 9.20, 10.57, and 7.38, respectively, and no significant (p>0.05) differences were observed among the treatment groups. Numbers of anovulated follicles were significantly (p<0.05) higher in pFSH-8 (6.20) than in pFSH-3 (0.20) and pFSH-5 (3.50). Total ovarian stimulation was increased linearly with increasing the dosage of pFSH and was highest in pFSH-8 (12.10) followed by pFSH-5 (10.90) and pFSH-3 (9.40). Out of ten, five donor does of pFSH-3 had no stimulation and only one donor does had anovulatory follicles. Three donor does in pFSH-5 and two donor does in pFSH-8 had no CL. Donor does with CL were only used in the result analysis.
Embryo production performances as affected by different dosages of pFSH are presented in Table 2. The number of CL was increased from pFSH-3 to pFSH-5 and then decreased from pFSH-5 to pFSH-8, but CL percentages ([no. of CL/total no. stimulation]×100) showed an inverse relationship with the increase of pFSH dosage, and the values were 97.87%, 67.89%, and 48.76% for pFSH-3, pFSH-5, and pFSH-8, respectively. Although the average number of CL per doe for the three treated groups were similar, the total number of recovered embryos and transferable embryos per doe were significantly (p<0.05) higher in pFSH-5 (7.00 and 6.71) than in pFSH-3 (3.00 and 2.80) and in pFSH-8 (2.00 and 1.50), respectively (Figure 1). The number of structures (embryos plus unfertilized oocytes) recovered and the recovery rate per doe were higher in pFSH-5 (7.14 and 69.22) followed by pFSH-8 (4.25 and 60.74) and pFSH-3 (4.00 and 59.73), respectively (Table 2). The number of unfertilized oocytes per doe was lower in pFSH-5 (0.14) followed by pFSH-3 (1.00) and pFSH-8 (2.25), (Figure 2), but no differences (p>0.05) were observed among treatment groups.
DISCUSSION
In this study, all the donor does of pFSH-5 and pFSH-8 showed the signs of estrus by using 14-day CIDR treatment. This value is similar to the findings of earlier researchers (Lehloenya et al., 2008; Lehloenya and Greyling, 2009) who recorded 100% estrus for Boer goats using 17 days CIDR and superovulated with pFSH. On the other hand, the present result was superior to some researchers who observed 87% and 75% estrus for Toggenburg goats (Fonseca et al., 2005; Zambrini, 2006). On the contrary, only 50% of does of pFSH-3 showed estrus in this study, and this result might be due to low dosage of pFSH, which may not be able to stimulate the ovary. These data indicate that CIDR with a sufficient amount of pFSH is an effective means of estrus synchronization for Boer crossbred does.
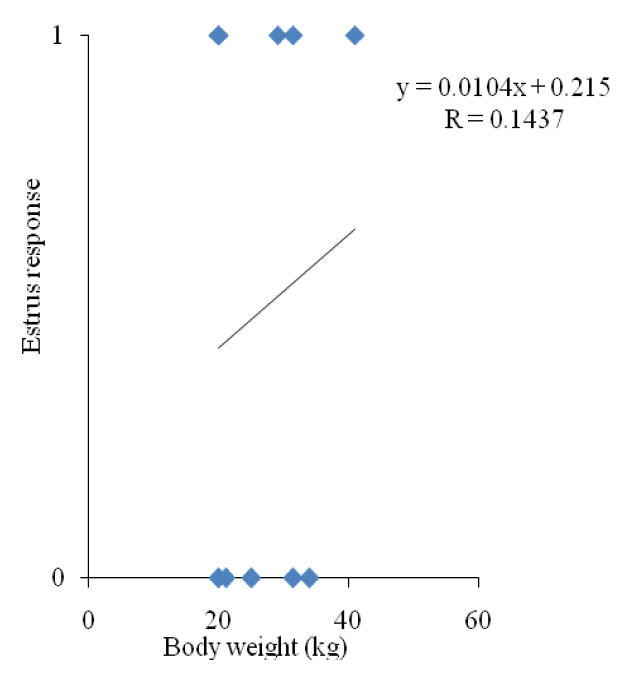
Although ovulation rate was similar for all the groups, pFSH-5 produced significantly a higher number of embryos than other groups. However, ovulation rate was not represented by embryos, which also included the ovulated oocytes that failed to be fertilized. It implies that the superovulation effect was not boosted up by high and low dosages of exogenous pFSH. In superovulated does, reduced fertility occurred due to prolonged elevation in estrogen concentrations resulting from the large anovulatory follicles and the abnormal maturation of oocytes (Cameron et al., 1988; Kumar et al., 1991; Ryan et al., 1991). Moreover, high estradiol concentration around and/or beyond ovulation induced disorders on the later reproductive process of fertilization and embryo development (Veiga-Lopez et al., 2006). Results of the present study also are in line with the findings of Ryan et al. (1991). In this study, the numbers of total and transferable embryo recovered from pFSH-3 and pFSH-8 were lower than pFSH-5. This result indicated that exogenous pFSH could promote ovarian responses, but the best superovulation results could be obtained only with a certain pFSH dosage range. However, the superovulation effect is not promoted by too high (>5 mg per kg BW) and too low (<5 mg per kg BW) dosages of exogenous pFSH as suggested in this study. Too high dosage of exogenous pFSH reduced the number for embryo production by producing a higher number of anovulatory follicles, which might increase estrogen levels in the blood. Elevated estrogen level in blood impaired sperm transportation in the female reproductive system and the breakage of sperm cells, which resulted in a higher number of unfertilized oocytes (Conley and Hawk, 1970; Armstrong et al., 1983; Greve et al., 1995). On the other hand, a too low dosage of exogenous pFSH may not be sufficient for stimulation of folliculogenesis and maturation of follicles. In this study, 50% of animal showed stimulation, which might be due to coincidence with the internal FSH production and the exogenous pFSH treatment. For pFSH-3, estrus response was not correlated with the BW (Figure 3).
The highest numbers of total embryo recovery, transferable embryo and structured recovered were obtained by using 5 mg pFSH per kg BW. These results were higher than the findings of previous studies (Menchaca et al., 2007; Abdullah et al., 2012; Xiao et al., 2013) but lower than the findings of an earlier study (Lehloenya, 2013). This result might be explained by the influence of breed, BW and nutrition (Bindon et al., 1986; Mani et al., 1992; Gonzalez-Bulnes et al., 2004; Abecia et al., 2006; Scaramuzzi et al., 2006).
Anovulatory follicles are obvious in the findings in any superovulatory protocols (Veiga-Lopez et al., 2006). In this study, the incidence was 22% to 45%, which was similar to the findings of previous studies (Gonzalez-Bulnes et al., 2000; Abdullah et al., 2012). The anovulatory follicle number was increased with the increasing dosage of FSH and the highest number obtained in this study was similar with the findings of a previous study (Abdullah et al., 2012). For pFSH-3, pFSH-5 and pFSH-8, the number of goat had anovulated follicle was 1, 3, and 6, respectively. This number was not enough to carry out correlation analysis between BW and anovulatory follicle. Therefore future studies involving dosages per kg BW need to establish this relationship. The causal factor for anovulation may be the presence of LH in the FSH preparation, which alters the endocrine and ovarian function after administration of a high dosage of exogenous gonadotrophin, and results in a smaller size and defective maturation of follicles (Murphy et al., 1984; Noel et al., 1994; D’Occhio et al., 1999; Kendall et al., 2004; Veiga-Lopez et al., 2006).
The dosage of pFSH had a significant effect on the embryo production performance. Five mg of pFSH per kg BW produced a significantly higher number of total and transferable embryos with the highest recovery rate. At the same time, this dosage produced the lowest number of degenerated embryos and unfertilized oocytes, but no differences were observed among the treatment groups. Results indicated that 5 mg pFSH per kg BW is suitable for producing embryos in the crossbred goats; however, further studies are needed to evaluate the interaction of FSH dosage with other factors (e.g., nutrition, age, surgery cycle) affecting superovulation performance in goats.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







