



INTRODUCTION
Japanese Black (JB) cattle are known for their highly marbled beef, greater content of unsaturated fatty acids (USFA), and lower melting point of fat compared to other breeds (Smith et al., 2006). In this breed, single-nucleotide polymorphisms (SNPs) in exon 5 of the GH gene have been reported to cause the amino acid substitution from CTG (leucine; Leu) to GTG (valine; Val) and from ACG (threonine; Thr) to ATG (methionine; Met) at codon 127 and 172, respectively. Moreover, there are 3 haplotypes observed in these codons, namely haplotype A (Leu-Thr), haplotype B (Val-Thr), and haplotype C (Val-Met) resulting in 6 genotypes, i.e., AA, AB, BB, AC, BC, and CC (Chikuni et al., 1997). Our previous studies (Katoh et al., 2008; Ardiyanti et al., 2009a; Ardiyanti et al., 2009b) have demonstrated that this polymorphism was related to characteristics in beef quality, carcass weights, and plasma hormone concentrations in JB cattle. Regarding i.m. fatty acid (FA) composition, JB cattle with haplotype A had greater carcass weight yet lower USFA contents, whereas those with haplotype C had lower carcass weight yet greater USFA contents.
Our present study was focused to analyze fatty acid synthesis in cattle with GH haplotypes A, B, and C. Three main enzymes that are involved in biosynthesis of lipid, i.e. fatty acid synthase (FASN), stearoyl-coA desaturase (SCD), and sterol regulatory element binding proteins-1 (SREBP-1), were selected for gene expression analysis. FASN and SCD are known for their roles in synthesizing saturated and unsaturated fatty acids, respectively (Ntambi, 1995; Chirala and Wakil, 2004). These genes are activated by SREBPs that bind to the sterol regulatory element sequences at their promoter regions (Shimano, 2001). A study using pig adipocytes has shown that GH decreases FASN mRNA level through reduction of FASN gene transcription and FASN mRNA stability. Previous reports on Holstein cows showed that GH reduces SCD mRNA levels in adipose tissue (Beswick and Kennelly, 2000). Insulin, on the other hand, is known to have stimulatory effects on SREBP-1, FASN, and SCD gene expression in adipose tissues (Yin et al., 1998; Shimano, 2001; Daniel et al., 2004).
Therefore, we aimed to determine if bovine GH gene polymorphism affects the lipogenesis regulation in 31-month old heifers by altering the lipogenic gene expressions.
MATERIALS AND METHODS
Animals
JB heifers (n = 111; 30.9┬▒0.9 months of age) raised under the same housing and feeding condition in Yamagata prefecture, Japan were used in this study. For hormones and lipogenic genes expression studies, heifers were grouped based on their GH genotypes, i.e. AA, AB, BB, AC, BC, and CC. For PearsonŌĆÖs correlation study, the heifers were divided into three haplotype groups according to presence of the related haplotypes, i.e., group haplotype A, B, or C.
Blood plasma and diaphragm tissues collection
Approximately 50 ml of whole blood sampled before slaughter was centrifuged at 2,000├Śg for 15 min at 4┬░C for plasma collection. Then, 200 U of heparin and 1 unit of trypsin inhibitor (Roche Applied Science, Indianapolis, USA) were added and the plasma was kept frozen at ŌłÆ30┬░C until analysis. For genomic DNA and total RNA isolation, diaphragm tissues were collected immediately after sacrifice, placed in RNase free vials, and kept frozen in liquid nitrogen before storage at ŌłÆ80┬░C.
Genomic DNA isolation and GH genotyping
Genomic DNA was isolated from approximately 50 mg of homogenized bovine diaphragm tissue using the QuickGene-800 automatic nucleic acid isolation system (Fujifilm, Tokyo, Japan). The eluted DNA concentration was measured using Nanodrop® ND-1000 Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA). The GH genotype was determined by allele-specific multiplex (ASM)-PCR according to the method of Chikuni et al. (1997) with a slight modification. ASM-PCR was carried out in 15-μ reaction mixtures containing 150 ng of template DNA, 10 mM Tris HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 200 μM each of dNTPs, 0.5 units of AmpliTaq Gold DNA polymerase (Applied Biosystems, Foster City, CA, USA), 10 pmol each of common primers GH4F and GH5R, and 5 pmol and 3 pmol of specific primers GHAR and GHABR (Sigma Genosys, Ishikari, Japan), respectively. The reverse primer GHAR, GHABR, and GH5R were designed to determine the GH haplotype A, B, and C, respectively. The sequences of primers are described in Table 1. Template DNA was amplified using a PC808 thermal cycler (Astec, Fukuoka, Japan) as follows: a primary cycle of denaturation (94°C for 9 min), 38 cycles of denaturation (94°C for 30 s), annealing (55°C for 20 s), and extension (72°C for 15 s), followed by one cycle of final extension (72°C for 7 min). The PCR products were then analyzed by horizontal electrophoresis (100 V for 35 min) through 2% agarose gel (Invitrogen, Carlsbad, CA, USA), ethidium bromide staining (0.5 μg/ml) for 40 min, and visualization by UV fluorescence using the Chemiimager digital imaging system (Alpha Innotech Corporation, San Diego, CA, USA). Single 347, 483, and 656-bp fragments were produced in GH genotypes AA, BB, and CC, respectively, whereas the combinations of two fragments with different sizes were produced in GH genotypes AB, AC, and BC.
Total RNA isolation and cDNA synthesis
Total RNA was isolated from approximately 100 mg of homogenized bovine diaphragm tissue using 1 ml of ISOGEN (Nippon Gene, Toyama, Japan). Before proceeding to cDNA synthesis, 5 μg of total RNA was incubated at 65°C for 10 min with DNase I (Invitrogen, Carlsbad, CA, USA). The DNase treated total RNA (0.5 μg) was then subjected to reverse transcriptase (RT) PCR for cDNA synthesis using High Capacity RNA-to-cDNA Master Mix (Applied Biosystem, Foster City, CA, USA) contaning both random primers and oligo(dT) primer. The RT-PCR was carried out in 96-well GeneAmp® PCR System 9700 (Applied Biosystem, Foster City, CA, USA) with condition as follows: 25°C for 5 min, 42°C for 30 min, and 85°C for 5 min.
Real-time PCR
Real-time PCR was conducted using Power SYBR┬« Green PCR Master Mix (Applied Biosystem, Foster City, CA, USA) and 7500 Real-Time PCR System (Applied Biosystem, Foster City, CA, USA). Sequence of primers for FASN, SCD, and SREBP-1 genes is listed in Table 2. Also, GAPDH (Taqman MGB P/N 4324034, Applied Biosystem) was used as the housekeeping gene to normalize the FASN, SCD, and SREBP-1 mRNA levels. The real-time PCR condition was performed with initial denaturation at 50┬░C for 2 min and 95┬░C for 10 min and 55 cycles of amplification at 95┬░C for 15 s and 60┬░C for 1 min. To analyze the differences of FASN, SCD, and SREBP-1 mRNA levels among genotypes and their correlations with fatty acid compositions, 2ŌłÆ╬ö╬öCt and 2ŌłÆ╬öCt were used, respectively.
Measurements of plasma GH, IGF-I, and insulin concentrations
The double-antibody RIA method was performed to measure GH and IGF-I concentrations, whereas insulin concentrations were measured using the single-antibody RIA method, as previously described by (Kuhara et al., 1991). Intra-assay coefficients and minimum detectable concentrations for GH, IGF-I, and insulin were 4.26% and 0.0603 ng/ml; 9.21% and 50 pg/ml; and 24.67% and 0.1008 ╬╝U/ml, respectively.
Gas chromatography
Ten milliliters of chloroform and methanol solution (2:1 v/v) was added to diaphragm tissues. The chloroform and methanol fractions were then separated by adding diluted sulphuric acid. Using rotary evaporator, the chloroform fraction was dried up at 60┬░C and the isolated lipid was diluted using hexane. Then, 0.5 N sodium methoxide-methanol was added to the mixture and the mixture was incubated at 80┬░C for 20 min for methyl esterification. The mixture was diluted with hexane and water and subjected to gas chromatography. The methylated lipid was analyzed using a gas chromatography (Hitachi G-3000, Tokyo, Japan) equipped with a 1.5 m├Ś3 mm glass column (Unisole-3000; GL Science Inc., Tokyo, Japan). The carrier gas was nitrogen and the column oven condition was programmed from an initial temperature of 205┬░C to a final temperature of 222┬░C at the rate of 1┬░C/min with the column head pressurized at 1.0 kg/cm2. Identification of fatty acids (myristic acid, C14:0; palmitic acid, C16:0; palmitoleic acid, C16:1; stearic acid, C18:0; oleic acid, C18:1; linoleic acid, C18:2) was achieved by comparing the relative retention time of fatty acid peaks from samples with those of standards. The amount of fatty acid contents was determined using its area percentage.
Statistical analysis
All data for hormone and gene expression measurements in genotype groups were expressed as the mean┬▒SE. Duncan s multiple range test was used to analyze the difference among genotype groups after one-way ANOVA (Excel Toukei, add-in software for MS- Excel, ESUMI Co., Ltd.). To analyze correlations between gene expression and fatty acid composition, PearsonŌĆÖs correlation coefficient between two variables was used. Differences were considered significant at p<0.05 and differences at p<0.10 were considered as tendencies.
RESULTS
Plasma GH, IGF-I, and insulin concentrations
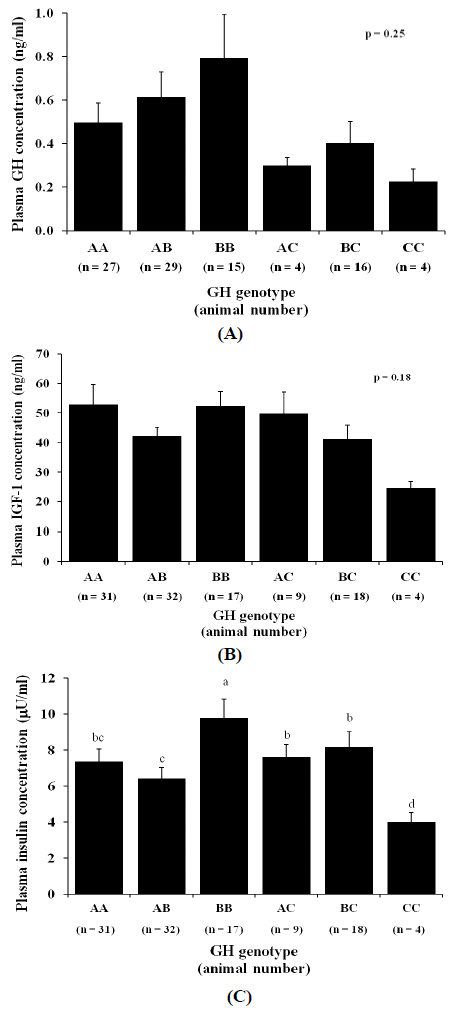
To observe the SNP effects on concentrations of hormones involved in somatotropic axis, we measured the plasma concentrations of GH, IGF-1, and insulin in heifers with different GH genotypes. As shown in Figure 1A and 1B, GH genotypes did not significantly alter plasma GH and IGF-1 concentrations (p = 0.25 and p = 0.18, respectively). However, for insulin concentrations, heifers with genotype CC had the lowest concentrations among all genotypes (Figure 1C; p = 0.02).
Genes expression levels of FASN, SCD, and SREBP-1
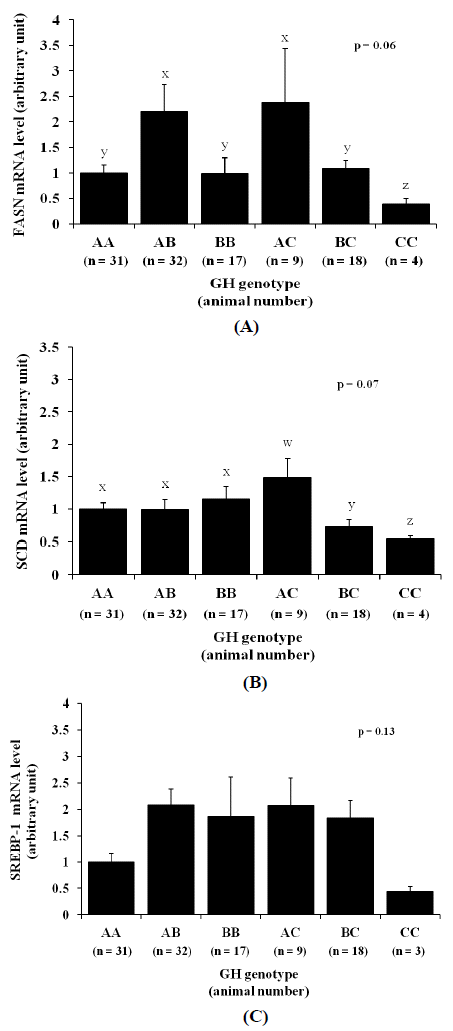
Next, we examined the SNP effects on gene expression levels of FASN, SCD, and SREBP-1 in diaphragm tissues. As demonstrated in Figure 2A and 2B, the expression levels of FASN and SCD genes in heifers with genotype CC tended to be the lowest among genotypes (p = 0.06 and p = 0.07, respectively) being approximately 50% lower than those in genotype AA. The lowest SREBP-1 mRNA level was also observed in heifers with genotype CC, although the difference was not statistically significant (Figure 2C; p = 0.13).
Correlations between lipogenic genes expressions levels and i.m. fatty acid compositions
Using the same PearsonŌĆÖs analysis, we examined if the haplotype differences affect the relationships between the lipogenic gene expression levels and i.m. fatty acid composition in diaphragm tissues (Table 3). However, due to a small population number of heifers with genotype CC (only 4 heifers out of 111 heifers), we grouped the heifers into group haplotype A, i.e. those with genotype AA, AB, and AC; haplotype B, i.e. genotype BB, AB, and BC; and haplotype C, i.e. genotype CC, AC, and BC. Although only at tendency levels, FASN mRNA levels in haplotype A positively correlated with SFA contents and negatively correlated with C18:2 and USFA contents. FASN mRNA level in haplotype B correlated with C16:0 content at tendency level and with C18:2 contents. However, FASN mRNA level in haplotype C did not show any correlations with the FA compositions. Moreover, SCD mRNA levels in haplotype A positively correlated with MUFA contents and negatively correlated with C18:0 content (p<0.05). The SCD mRNA levels in haplotype A also tended to positively correlate with C16:1, C18:1, and USFA contents and USFA/SFA ratio while negatively correlate with SFA content (p<0.10). SCD mRNA levels in haplotypes B and C, however, did not correlate with any of the FA compositions. As for SREBP-1, there was no correlation found between SREBP-1 mRNA levels and the FA compositions regardless of the haplotypes.
DISCUSSION
Based on our previous studies (Ardiyanti et al., 2009a; Ardiyanti et al., 2009b) on GH gene polymorphism in JB cattle, our present study addressed the GH SNP effects on somatotropic hormone concentrations and lipogenesis in JB heifers. We previously have demonstrated that 10 mo-old heifers with genotype AC, BC, and CC had significantly greater plasma GH concentrations than those with other genotypes (Ardiyanti et al., 2009a). This result led us to speculate that the SNP might alter GH-GHR binding efficiency followed by a GH resistance condition and hence increased GH release. However, in our present study, basal GH and IGF-I concentrations did not significantly differ among GH genotypes. It is possible that the absence of the SNP effects on GH concentrations in the present study was due to animal age difference. Heifers used in this study were 31 months of age, during which GH concentrations were probably already declining and thus dampening GH gene polymorphism effects. Subsequently, it was not surprising when the plasma IGF-1 was found at statistically same concentrations among genotypes. However, it is noteworthy that we only did a single measurement for the plasma GH concentration, which probably could not represent the overall effect of the SNP on the pulsatile release of GH. Plasma insulin was found at the lowest concentrations in animals with genotype CC and this result was in accordance with our previous results (Ardiyanti et al., 2009a). GH excess is known to impair cell sensitivity to insulin resulting in insulin resistance. Moreover, GH administration has been linked to increased insulin release from the pancreas through STAT5A signaling (Etherton and Bauman, 1998; Dominici et al., 2005; Feng et al., 2009).
In regards to lipogenic gene expressions, it is surprising that despite the lower SFA and the greater USFA contents shown by cattle with genotype CC in previous studies, our present results revealed that FASN and SCD mRNA levels in heifers with genotype CC were in fact the lowest among genotypes. However, SREBP-1 mRNA level was not affected by the genotype difference indicating that the SNPŌĆÖs effects on FASN and SCD mRNA levels might be independent from SREBP-1. Also, given that the promoter regions of FASN and SCD genes are activated by insulin (Latasa et al., 2000; Keating et al., 2006), the low level of insulin concentration in heifers with genotype CC might relate to their low FASN and SCD mRNA levels as well.
As reviewed in another study (Shimano, 2001), FASN and SCD genes are among other genes activated by SREBP-1 and therefore it is very likely that FASN and SCD mRNA levels would be simultaneously activated. Our correlation analysis has indeed demonstrated strong positive correlations among FASN, SCD and SREBP-1 mRNA, however, the haplotype differences did not seem to alter the correlations, except for haplotype B where FASN mRNA level did not correlate with SREBP-1 mRNA level (data not shown). This result suggests that the GH SNP alters the FASN and SCD expression levels without involving SREBP-1.
Our analysis of the FA compositions, however, has shown interesting results where the correlations between FASN and SCD mRNA levels with the FA contents were more pronounced in group haplotype A than in haplotypes B and C. FASN and SCD are known for their roles in synthesizing saturated and unsaturated fatty acids, respectively (Ntambi, 1995; Chirala and Wakil, 2004). This is accordingly reflected in group haplotype A through the positive correlations between FASN mRNA levels and SFA contents and the negative correlations between SCD mRNA levels and USFA contents. In contrast, there was no correlation between the gene expression levels and FA compositions observed in haplotype C. Moreover, the only correlations shown in haplotype B were between FASN mRNA levels and C16:0 and C18:2 contents. These results implied that the GH SNP has not only altered the gene expression levels of FASN and SCD, but also the relationships between the gene expressions of those enzymes and their FA products.
GH treatment has been previously shown to reduce SCD gene expression to undetectable levels in bovine adipose tissues and this was considered to be the direct effect of GH on SCD (Beswick and Kennelly, 2000). In different studies, GH has also been shown to negate the insulin effect to induce FASN and SCD transcriptions in rats and pigs (Yin et al., 1998). Moreover, STAT5A also has been shown to directly repress FASN mRNA levels in 3T3-L1 cells (Hogan and Stephens, 2005). We have speculated on the hypothetical alteration of GH-GHR binding efficiency in GH haplotype C that might lessen the repression effect by STAT5 on promoter regions of FASN and SCD genes. However, our present study has shown conflicting results where the mRNA levels of both enzymes were lower in genotype CC, although this might be attributed to the low insulin concentrations and tissue differences. On the other hand, SREBP-1 mRNA in porcine adipocyte was shown to be unaffected by GH or insulin or both treatment (Louveau and Gondret, 2004). These studies might support our result where GH haplotype difference did not affect the SREBP mRNA levels or its relationship with any of the FA contents either. Therefore, we propose that the alteration of FASN and SCD mRNA levels and FA synthesis by the SNP in GH gene involves direct action of insulin and is independent from SREBP-1.
In our present study, we demonstrated the effects of GH gene polymorphism on lipogenic genes expression levels in diaphragm tissues of JB heifers at 31 months of age. The variation of GH haplotypes altered the regulation of FASN and SCD gene expressions and fatty acid synthesis in the tissues as well. However, it is a limitation in our study that we measured the lipogenic gene expression levels in diaphragm tissues instead of adipose tissues, which is the main lipogenesis site in ruminants. The haplotype grouping method we used based on the presence of haplotype A, B, and C due to the small number of heifers with genotype CC is also another weakness that might interfere with the relationship of the gene expressions with FA compositions. Therefore, further studies are required to elucidate the detailed mechanism on how the GH gene polymorphism affects the lipogenesis mechanism in cattle.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







