



INTRODUCTION
In tropical zones, where day (light hours) variation is minimum, sheep are not seasonal, in contrast to breeds from other latitudes where annual variation in day length is greater (Martinez et al., 2001). Nevertheless, factors such as high temperatures and lack of feed can limit their sexual activity for some months, mainly the first months of the year (Castillo et al., 1972; Cruz et al., 1994). Several studies have proven that nutrition constitutes one of the factors that greatly influence the reproductive processes of male and female domestic animals (Martin and Walkden-Brown, 1995; Leroy et al., 2005). Supplementing the ewes with nutritious feed before mating, known as flushing, has been used to increase ovulation ratio and embryo survival (Johnson et al., 1990). The effects of nutrition on follicle development and ovulation ratio can potentially be mediated by changes in the circulation of metabolites (glucose, amino acids, etc.), gonadotropins (LH and FSH), metabolic hormones, or a combination of these factors (Rhind, 1992). To make up for the protein deficiency of tropical grasses (5 to 7%), some species of aquatic plants have been used as feed supplement for ruminants (Gallego et al., 1994). Duckweed contains 20 to 25% protein, which can be increased, through the use of organic fertilizers, up to 45%, and thus can be utilized to increase the nutritional value of the diet (Huque et al., 1996; Cheng and Stomp, 2009), since it is highly digestible given its low lignin content (Castillo et al., 2005). On the other hand, Taiwan grass (Pennisetum purpureum) is one of the most important tropical forages and is widely spread in tropical and subtropical regions of the world (Zhang et al., 2011). The objective of this work was to determine the effect of duckweed supplementation in Taiwan grass hay based diets on some reproductive variables of crossbred sheep from Pelibuey, Dorper, and Katahdin breeds.
MATERIALS AND METHODS
Experimental site, animals, and synchronization protocol
In this study, 18 multiparous, crossbred ewes from Pelibuey, Dorper, and Katahdin breeds were used (39.7±4.0 kg), with a body condition from 3 to 3.5 in a 1 to 5 scale (Cottle, 1991), and similar characteristics regarding size and age. They were kept in individual pens, with free access to water and mineral salts. Before beginning the experiment, they were dewormed and vaccinated. The ewes were synchronized with intravaginal sponges impregnated with 40 mg fluorogestone acetate (FGA, ChronogestTM) and an intramuscular application of 400 UI equine chorionic gonadotropin (eCG, FolligonTM) at the moment of removing the sponges, which were placed 94 h after supplying the diets and removed 12 d later.
Experimental diets, chemical analysis and experimental design
Taiwan grass hay (TWH) (50 d after regrowth) used as a base diet, was supplemented with sundried duckweed (DW) (Lemna sp. and Spirodela sp.) to improve the protein content of the diet. DW was fed at 7:00 h, and TWH was given ad libitum at 8:00 and 16:00 h. Ewes were randomly assigned to the following treatments (n = 6): T1: TWH (control), T2: TWH plus 200 g DW, T3: TWH plus 300 g DW.
TWH and DW were analyzed for dry matter (DM), organic matter (OM) and crude protein (CP) according to the AOAC (2005). Neutral detergent fiber (NDF) and acid detergent fiber (ADF) by Van Soest et al. (1991).
Evaluated variables
Dry matter, crude protein intake and progesterone concentration in plasma
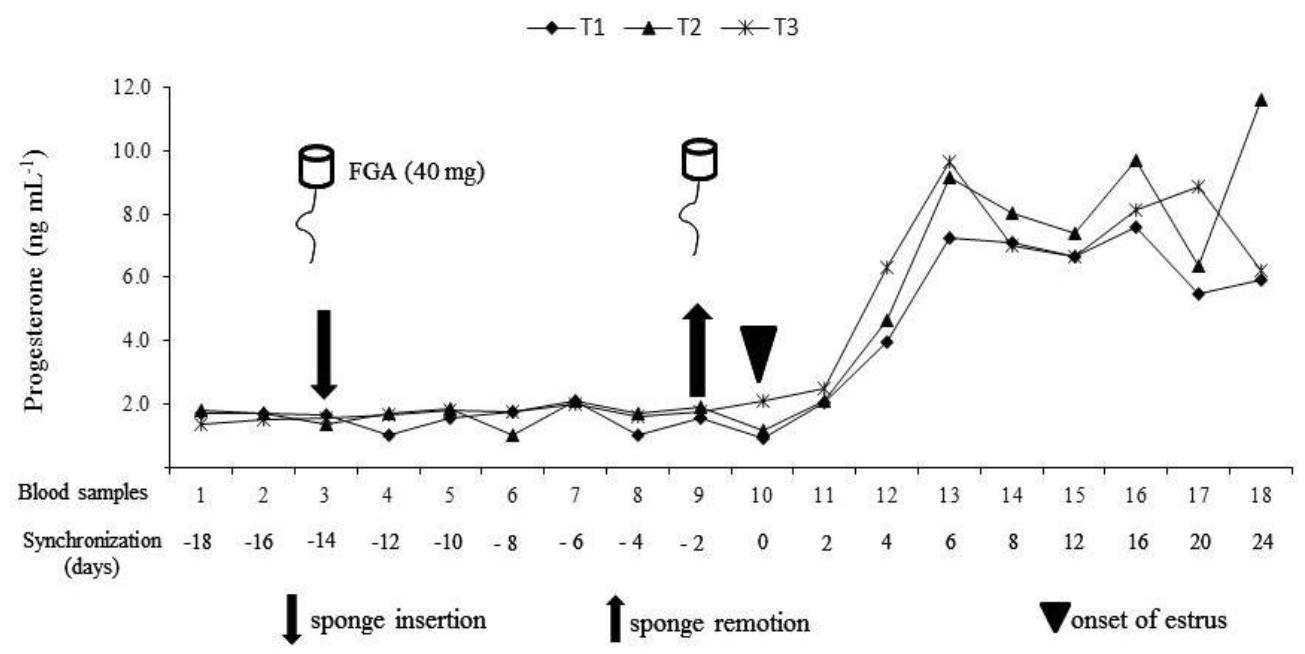
Dry matter intake (DMI) and crude protein intake (CPI) were measured through the difference between offered and rejected DM or CP (g animâ1 dâ1). To determine the progesterone concentration (P4), 5 ml of blood were individually collected through jugular puncture, centrifuged at 3,000 rpm for 15 min and the plasma obtained was kept at â4°C. The first and second samples were collected 94 h and 48 h, respectively, before synchronization, the third sampling at the moment of inserting the sponges, and then every third day. The last four samples were collected every four days, with a total of 18 samplings (Figure 1). To quantify P4, the tube enzymatic immunoassay (EIA) methodology (Reimers, 1984) was used.
Estrus detection, ewes repeating estrus and pregnancy diagnosis
Estrus detection began 12 h after removing the sponges and every 2 h for 20 min, using 4 rams with aprons. The beginning was considered from the moment the ewe allowed the ram to mate, finalizing when it was no longer receptive. The ewes were mated twice, at the beginning of estrus and 12 h later. The rams were rotated every four services to avoid exhaustion and allow semen production.
To detect the return to estrus, 14 d after the last day of mating, the ewes were exposed to 4 rams, considering that the sexual cycle of sheep is 17±3 d. The same procedure was used to detect estrus. The pregnancy diagnosis was done 21 d after the last day of mating with the animal standing and using a transabdominal technique with a frequency of 3.0 MHz (Memon and Ott, 1979). A Digital Veterinary Ultrasound Imaging System, model CTS-3300V, SIUI brand equipment was used.
Statistical analysis
Data on DMI, CPI, onset and beginning of estrus, were analyzed as a completely randomized design using the GLM procedure (SAS, 2000). Initial weight was used as a covariate for DMI and CPI. Data on P4 was analyzed using the MIXED procedure of SAS. Percentage of estrus, ewes that repeated estrus, and rate of pregnant ewes were analyzed using Chi-square test and Fisher exact test.
RESULTS AND DISCUSSION
Chemical composition of forages
Table 1 shows the chemical composition of TWH and DW. The results agree with those reported by Kozloski et al. (2006) and Da Silva et al. (2007) regarding TWH; however, CP content differs from the results obtained by Tessena and Baars (2004). This could be due to type of fertilization used on the graminea. Regarding DW, the values obtained coincide with those found by Huque et al. (1996) and Olorunfemi et al. (2006).
Dry matter and crude protein intake
DMI was similar for all three treatments (p>0.05) (Table 2); although there was a slight decrease in the intake of the base diet as DW was supplemented. Similar results were obtained by Aregheore (2006) who supplemented copra cake in Taiwan grass diets. Kahindi et al. (2007) reported a decrease in Taiwan grass intake when supplementation with Madras thorn (Pithecellobium dulce) increased. Abdulrazak et al. (1996) observed a decrease in the intake of Taiwan grass when supplemented with increasing levels of gliricidia (Gliricidia sepium), which they attributed to legumeâs volume. Supplementation with DW caused a slight decrease in the intake of TWH, although total DMI among treatments was not different (p>0.05); however, this could be justified by the restriction of DW, which was not fed ad libitum. On the other hand, fertilization of the DW with ovine manure could have affected taste and smell, which in turn decreased DMI, and probably the presence of antinutritional factors in DW (Bairagi et al., 2002; Kalita et al., 2007). Supplementation with DW increased intake of CP (p<0.05), where T3 was different from T2 and T1 (Table 2). An increase in the intake of CP was observed when supplementing with 300 g DW (p<0.01), with no difference between T1 and T2. Daily CP intake in all three treatments was higher than the recommended by NRC (1985).
Onset, beginning and duration of estrus
Table 3 shows the beginning of estrus after removing the synchronization device. No difference was found on the beginning of estrus (p>0.05) (35.72, 34.83, and 42.95 h for T1, T2, and T3, respectively). These results coincide with Molina et al. (2005), who reported an interval in the onset of estrus of 32 to 36 h, but differ from Camacho-Ronquillo et al. (2008), who observed a beginning of estrus between 21.4 and 24.2 h, and attribute the early appearance of estrus to the application of eCG two days before removing the FGA sponges. In the present study, eCG was applied at the moment of removing the sponges and most estrus appeared in the first 48 h, which coincides with Mendoza (2008) and Fukuy et al. (1999) when using a similar synchronization protocol.
DW supplementation did not affect the percentage of estrus appearance (p>0.05) (Table 3). In T1 and T2, one ewe did not respond to the synchronization protocol; however, they were not different (p>0.05) from T3. A deficient P4 absorption from the intravaginal sponges could have been the cause of this lack of response to synchronization (Kenji et al., 2004); when the device with P4 was inserted, the ewes were at different stages of the oestral cycle, which could have caused a variation in the duration of endogenous P4 secretion. According to Leyva et al. (1998), the changes resulting from the combination of exogenous and endogenous P4 can cause alterations in the follicular dynamics and variations in the appearance of estrus, once the device is removed. The variations in the ovarian response seem to be due to the differences in the degree of follicular maturation, presence of ovulatory follicles, non ovulatory follicles, and subfunctional corpora lutea (Liu et al., 2007).
DW supplementation had no effect on the number of ewes presenting estrus, nor on the duration of these (p>0.05) (Table 3). Gordon (1997) and Fernandez et al. (1997) point out that estrus duration in sheep varies from 24 to 48 h, can increase by up to 50% in ewes with multiple ovulations (Camacho-Ronquillo et al., 2008), or decrease by half when the ewes are kept with the ram, compared to ewes where estrus is detected with the ram every 2 h (Gonzalez et al., 2000). The results obtained in the appearance of estrus (Table 3) were within the 24 to 48 h, and coincide with Mazzarri et al. (1976), but differ from Cordova-Izquierdo et al. (1999) who mention a 26 h duration, and attribute it to the period of non reproductive season, since in the season of ovarian cyclicity the duration can regularly reach 30 h. Nevertheless, Camacho-Ronquillo et al. (2008) report estrus duration of up to 60 h, which they attribute to a greater number of ovulatory follicles, from the effect of applying eCG two days before removing the progestagen. Navarrete-Sierra et al. (2008) used diets with high quality protein in ewes synchronized with FGA for superovulatory treatment based on ovine follicle stimulating hormone, and observed an increase in the ovulation rate and secretion of P4, as well as an extension of estrus through the lengthening of the midcycle lifespan of the corpus luteum.
Percentage of ewes repeating estrus
Using the same protocol to detect estrus, no ewe returned to estrus. Lozano et al. (2003) mention that low energy diets during the embryo development stage increases in vitro production of PGF2α; which could cause luteolisis and embryonic death, which causes a return to estrous cycles from the inability of maintaining gestation. Martinez (1999) reports that 89.4% of the ewes did not return to estrus (17 out of 19 animals), and attributes it to the fact that the sanitary and nutritional conditions were adequate. Mendoza (2008) points out that only one ewe returned to estrus, and mentions that this was due to the fact that the nutrition supplied was adequate. DW supplementation did not affect the return to estrus, which suggests that the ewes were able to obtain enough energy and protein from the diet to sustain gestation and avoid return to estrus.
Pregnancy rate
DW supplementation had no influence (p>0.05) on gestation percentage (Table 3). The pregnancy diagnosis showed that ewes with T1 and T2 had 80% pregnancy, while those with T3 had 83%. Protein supplementation before mating, mainly with amino acids like lysine and methionine, can increase fertility, since several hormones (hypothalamic, pituitary, and ovarian) are of protein origin (Mitchell et al., 1998; Bell and Bauman, 2006). The explanation to the response obtained in this study could be due to the high rumen CP degradability of DW (Khan et al., 2002), which decreases the supply of amino acids; and although an increase in the intake of this nutrient stimulates the ovulation rate, the effect is more evident when the intake prior to flushing is at maintenance level or slightly below it (Fletcher, 1981). However, in ruminants, reproduction has been directly associated with energy availability, decreasing the ovulation rate when this nutrient is reduced in the diet (Wathes et al., 2007). Energy and protein supply influence the ovulation rate, but in order to reach the maximum effect, an increase of both nutrients may be needed (Fernandez et al., 2007). Energy or protein flushing does not always allow for homogeneous responses to exogenous stimuli of multiple ovulation (Lozano et al., 2003). The obtained results (81% mean in all 3 treatments) are similar to those reported by Camacho-Ronquillo et al. (2008), who observed 85.7% pregnancy with a diet based on oats and alfalfa, as well as a commercial concentrate with 16% protein, ad libitum.
Progesterone concentration
Supplementation did not affect P4 concentration (p>0.05). However, the sampling period was different (p<0.01), although the treatmentĂperiod interaction showed no differences (p>0.05). P4 concentrations increased significantly (p<0.01) starting on d 4 of the sampling (Figure 1), approximately 4 d after removing the sponge impregnated with FGA and the application of eCG, when the corpus luteum finished its formation and increase P4 secretion as it matures, since it is necessary to maintain sheep pregnancy (Bartlewski et al., 1999). This coincides with the reports by Uribe-Velasquez et al. (2008) and Mendoza (2008), who also synchronized with FGA and eCG. It is considered that circulating levels of P4 greater than 2 ng mlâ1 are indicative that the luteal tissue is active and there is a probable pregnancy (Restall et al., 1990); however, even if these levels are detected, some ewes do not reach lambing since they can develop pyometra, condition in which the corpus luteum remains active (Ortega, 1997), or because of embryonic death, which occurs in 2 to 8%, even up to 20%, of the cases under normal conditions (Fowler and Wilkins, 1984; Bretzlaff et al., 1993). P4 concentration in the blood for early diagnosis of pregnancy is a technique used 19 to 23 d post-service in ruminants (Ortega-Pacheco et al., 1999). High P4 levels suggest the presence of an active corpus luteum, which is compatible with pregnancy (Matsas, 1993). Under this premise, it is probable that all the ewes in heat and mated were pregnant in the last sampling (21 d post-service), considering that the P4 concentrations were above 2 ng mlâ1.
However, Gonzalez and Botero (1983) and Ortega-Pacheco et al. (1999) report a low efficiency of ultrasonography to detect pregnancy at 21 d, regardless of the technique used, since the size of the fetal sacks is too small, and detection may be hindered by diverse factors such as corporal condition, fasting, and experience of the operator, then they consider P4 concentrations in plasma a more reliable test. In this research, detection of pregnancy was performed 24 d post-service. The factors previously mentioned could explain that pregnancy was not detected in some ewes with a probability of presenting it, given that they had P4 concentrations above 2 ng mlâ1.
P4 concentration in the three samplings before synchronization (d â16) was different (p<0.05) to the P4 concentration obtained in the samplings collected on day 4 which was at the time when the corpus luteum was finishing its formation and day 21 when pregnancy was detected. However, it was not different (p>0.05) from the samplings corresponding to the beginning of estrus (d 0). P4 secretion can be affected by the reproductive stage of the ewes; however, the corpus luteum capability to secrete P4 is also a factor associated with P4 concentration in the blood. Evans and Robinson (1980) mention that it is possible to establish the existence of a direct relationship among eCG doses, corpus luteum, ovulation rate, and P4 concentrations in plasma in ewes previously treated with this progestagen.
CONCLUSIONS
The results obtained showed that DMI and CPI were not affected by DW supplementation. However, a slight decrease in TWH intake was observed as the DW supplementation levels increased. The number of ewes presenting estrus, the beginning and duration of estrus, pregnancy rate and P4 concentration in plasma were not affected by DW supplementation in diets based on TWH.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







