Effect of Sea Buckthorn Leaves on Inosine Monophosphate and Adenylosuccinatelyase Gene Expression in Broilers during Heat Stress
Article information
Abstract
The trial was conducted to evaluate the effects of sea buckthorn leaves (SBL) on meat flavor in broilers during heat stress. A total 360 one-day-old Arbor Acre (AA) broilers (male) were randomly allotted to 4 treatments with 6 replicates pens pretreatment and 15 birds per pen. The control group was fed a basal diet, the experimental group I, II and III were fed the basal diet supplemented with 0.25%, 0.5%, 1% SBL, respectively. During the 4th week, broilers were exposed to heat stress conditions (36±2°C), after which, muscle and liver samples were collected. High performance liquid chromatography (HPLC) was performed to measure the content of inosine monophosphate (IMP); Real-Time PCR was performed to determine the expression of the ADSL gene. The results showed that the content of breast muscle IMP of group I, II and III was significantly increased 68%, 102% and 103% (p<0.01) compared with the control, respectively; the content of thigh muscle IMP of group II and III was significantly increased 56% and 58% (p<0.01), respectively. Additionally, ADSL mRNA expression in group I, II and III was increased significantly 80%, 65% and 49% (p<0.01) compared with the control, respectively. The content of IMP and expression of ADSL mRNA were increased by basal diet supplemented with SBL, therefore, the decrease of meat flavor caused by heat stress was relieved.
INTRODUCTION
Sea buckthorn (Hippophae rhamnoides) is a small shrub comprised of fruit and leaves that are rich in nutrients and bioactive components such as vitamins (Kudritskaya et al., 1989; Zadernowski et al., 2003; Luhua et al., 2004; Ranjith et al., 2006), amino acids (Yushipitsina et al., 1988; Repyakh et al., 1990), lipids (Goncharova and Glushenkova, 1993; Ul’chenko et al., 1995; Bekker and Giushenkova, 1997), sugars and acids (Yang, 2009), and flavonoids (Häkkinen et al., 1999). Recently studies have shown that sea buckthorn has antioxidant (Geetha et al., 2002a; Geetha et al., 2002b; Chawla et al., 2007; Püssa et al., 2007; Geetha et al., 2009), anticancer (Olsson et al., 2004; Ma et al., 2007; Boivin et al., 2007), antiinfective (Larmo et al., 2008) and exerts beneficial effects on liver fibrosis (Gao et al., 2003) and immune function (Dorhoi et al., 2006). Sea buckthorn decreases the damage of enzyme activity or hormonal metabolic organism status caused by immobilization or cold-hypoxia-restraint (Krylova et al., 2000; Saggu and Kumar, 2007), however, its effects on heat stress is unclear. Heat stress is one of the most universal stress factors affecting poultry (Gregory, 2010), and is a major contributor to growth restriction (Liu, 2011; Melesse et al., 2011). In addition, heat stress can also influence the meat quality and flavor of poultry (Miao, 2007; Chiang et al., 2008; Lu et al., 2009).
Muscle inosine monophosphate (IMP) content is an indicator meat flavor and adenylosuccinatelyase (ADSL) is a key enzymes regulating IMP synthesis. Zhang et al. (2006) indicated there was positive correlation between the expression of ADSL gene and muscle IMP content. Therefore, the goal of the current study was to assess the effect of sea buckthorn leaves (SBL) on ADSL gene expression and IMP content were investigated during heat stress in poultry.
MATERIALS AND METHODS
Experimental birds, diets, and treatment
A total of 360 one-day-old male AA broilers (Harbin Delin broiler Co., Harbin, China) were randomly allotted to 4 groups of 90 birds each, with 6 replicates of 15 birds each. SBL that was collected in the Farm 8511 in June (Mishan, China) was shattered into powder with passing 40 screen mesh. The control group was fed a basal diet while the experimental group I, II and III were fed the basal diet supplemented with 0.25%, 0.5%, 1% SBL powder, respectively. Small amounts of the basal diet were first mixed with the respective amounts of SBL powder as a small batch and then mixed with a larger amount of the basal diet until the total amounts of the respective diets were homogenously mixed. The basal maize-soybean diet (Table 1) was formulated to refer to the NY/T 33-2004 (Chinese, 2004) and NRC (1994) nutrient requirement. Diets and fresh water were offered ad libitum. During the 4th week (from the 22th day to the 28th day), birds were exposed to heat stress at 36±2°C for 12 h/d (06:00 to 18:00) and 22°C for 12 h/d (18:00 to 06:00).
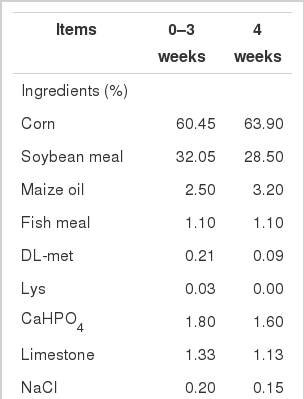
Composition and nutrient level of basal diet (air-dry basis)
Sampling time and procedures
At the 29th day, 2 birds per replicate were randomly killed after fasting 12 h. by cervical dislocation. The carcasses were then opened and the left breast and thigh muscle and liver samples were rapidly harvested and stored at −80°C until analysis. The Committee on Animal Care at Northeast Agricultural University approved the animal care procedures employed.
Assay of IMP content
Assay of IMP content in muscles
One gram of muscle sample was shredded and homogenated two times in chilled 10 ml 3.5% HClO4. The resulting homogenate was centrifuged at 3,500×g for 10 min and the supernatant was collected, the sediment was washed in 5 ml 3.5% HClO4, centrifuged and the supernatant collected and combined with the first supernatant. Double distilled water (ddH2O) was added at twice the volume of the supernatant up to 25 ml. The diluted supernatant was pipetted in 1 ml aliquots and 9 ml distilled water was added. Samples were filtered by 0.45 μm filter membrane and separated by high-performance liquid chromatography (HPLC) equipped with the 600E Multisolvent Delivery System and a UV 2487 Detector with Empower software (Waters Co., Milford, USA). The chromatographic column was Symmetry Shield™ RP18 (C18), 5 μm (Waters Co., Milford, USA). Mobile phase was phosphate triethyl ammonium and methanol (Sigma Co., USA) (95:5), all reagents were chromatographic pure; the flow rate was 1.0 ml/min; the detection wavelength was 254 nm; the chromatographic column temperature was 25°C; the sample size was 10 μl.
Preparation of standard curve
The IMP standard substance (Sigma chemicals, St. Louis, USA) was accurately weighed and then dissolved in the appropriate volume of Mobile phase to produce corresponding standard solutions at six concentrations in the range of 0.25 to 8 μg/ml (multiproportion). The IMP standard curves were plotted after linear regression of the peak areas versus concentrations.
Assay of gene expression
Assay of ADSL gene expression in liver
Total RNA was extracted from liver tissue, referring to the method offered by Shanghai Huashun RNArose kit (Huashun biotechnology Co., Shanghai, China). The RNA quality was detected by formaldehyde gel apomorphosis electrophoresis, the RNA concentration and purity was determined by spectrophotometer at A260/A280, and 2 μg total RNA was reverse transcribed into cDNA using M-MLV reverse transcriptase (Promega Co., Madison, USA). ADSL primers were synthesized by Shanghai biotechnology Co. (China), referring to the GeneBank cDNA sequence (Table 2), and the reference gene was 18sRNA (Yusuke Matsubara et al., 2005).

Primer sequence, product size and position
Amplicons via 1% agarose gel electrophoresis (AGE) chosen for sequencing were purified by using a TIANgel Midi Purification Kit (Tiangen biotechnology Co., Beijing, China) prior to sequencing. After purification, the PCR product was sequenced by Shanghai biotechnology Co. (China), and the sequencing primer sets were the same as those used for Real-Time PCR. Target genes were determined by Real-Time PCR after validating the sequence homologies.
Real-Time PCR procedure referred to SYBR® Premix Ex Taq™ kit instruction obtained from TaKaRa biotechnology Co. (Dalian, China), and 1 μl cDNA template was introduced into the reaction that was carried out in 20 μl ABI PRISM 7500 Real-Time PCR Reaction System (Applied Biosystems Co., USA). Temperature cycles were as follows: 95°C pre-degeneration 10 s, 95°C degeneration 5 s, 60°C renaturation/extension 34 s.
Preparation of amplification efficiency curve
The cDNA was gradient diluted according to Table 3, and various concentrations of cDNA were amplified by Real-Time PCR with the primers of ADSL and 18sRNA genes. The reaction system and procedure were in accordance to above. The amplification efficiency curves were plotted after linear regression of the cycle threshold (Ct value) versus Log 10diluion. Amplification efficiency E = 10−1/slope-1.
Statistical analyses
Data was analyzed by using one-factorial ANOVA procedure of SAS 8.0. Statistical significance was assumed at p<0.01. Data were expressed as mean±pooled SEM. Replicate was considered as the experimental unit. Number (n) used for statistics is note in the tables.
RESULTS
Effects of SBL on muscle IMP content in AA broilers
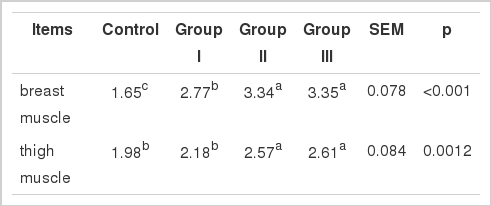
The effects of SBL on muscle IMP content are summarized in Table 3. Compared with the control, the content of breast muscle IMP of group I, II and III was significantly increased 68, 102, and 103% (p<0.01), respectively; the content of thigh muscle IMP of group II and III was significantly increased 56, and 58% (p<0.01), respectively (Table 4), but there was no significant influence in group I (p>0.05).

Effects of SBL on ADSL mRNA expression in liver in AA broilers
The regression equation of ADSL gene amplification curve is as follows: y = −3.2246x+32.776, R2 = 0.9972; the regression equation of 18sRNA gene amplification curve is as follows: y = −3.2025x+33.081, R2 = 0.9976. The amplification efficiency of the target gene is similar to that of reference gene from amplification efficiency E. The ADSL gene relative expression was expressed as 2−ΔCt (Feng, 2007), and its gene relative expression of group I, II and III were significantly increased 80, 65, and 49% (p<0.01), respectively, compared to chicks fed basal diet (Table 4).
DISCUSSION
Heat stress is one of the most universal and predominant factors that restrict the development of poultry in modern cultivation facilities. A number of bioactive components have been used to reduce heat stress, including tea polyphenols (Wang, 2004; Liu, 2005), extraction of fructus ligustri lucidi (Zhuang, 2007), Chinese magnoliavine fruit (Ma et al., 2005), lycopene (Sahin et al., 2006), and Turkish propolis (TatlıSeven et al., 2007). It has been reported that vitamin E, vitamin C and flavones also reduce the detrimental effects of heat stress (Sahin et al., 2002a; Sahin et al., 2002b; Chen, 2004; Wang, 2004; Huang, 2005). These are major constituents of sea buckthorn, however, there has been no direct study of the anti-heat stress of sea buckthorn. Heat stress not only decreases poultry performance (Chiang et al., 2008), but also the meat flavor. Meat flavor is influenced by flavor precursors, including amino acids, carbohydrates, inorganic salts and nucleotides. ATP is the major nucleotide in living muscle, however, during rigor mortis, it is converted into IMP. IMP is thought to be one of the most important factors in flavor development as it is a source of reducing sugar and nitrogenous base for Maillard reactions and acts as a flavor enhancer (Bayliss et al., 1995). Herein, SBL as a feed additive decreased the damage to meat flavor by increasing the muscle IMP content, but the interaction between IMP and other meat flavor precursors and its organoleptic properties remains to be determined.
One of the mechanisms of action whereby SBL increased IMP content was investigated. ADSL is an important enzyme that catalyzes two similar reactions in the de novo purine biosynthesis pathway. It catalyses the conversion of succinylaminoimidazole carboxamide ribotide (SAICAR) to aminoimidazole carboxamide ribotide (AICAR), as well as, the formation of adenosine monophosphate (AMP) from adenylosuccinate in the purine nucleotide cycle (Kmoch et al., 2000; Toth and Yeates, 2000). The content of IMP in muscle is influenced by pH, temperature and water activity (Vani et al., 2005; Kavitha et al., 2006), however, the background in raw meat is determined by ADSL gene expression (Zhang et al., 2006). All three SBL levels increased the ADSL gene expression. In a previous study, estrogen significantly increased the transcription rate of the ADSL gene (Zhu et al., 2001). The flavones in sea buckthorn are phytoestrogen that can exert estrogenic or anti-estrogenic actions depending on concentration, so it is possible that the SBL increased the ADSL gene expression in liver via its estrogenic activity. Generally, the SBL increased muscle IMP content by increasing ADSL gene expression, but the basal diet supplemented with 0.25% SBL didn’t increase the IMP content of thigh muscle although it increased ADSL gene expression, implying that dosage is an important factor. In addition, Kavitha and Modi (2006) found that the IMP content of fresh breast muscle was higher than that of fresh thigh muscle, suggesting that the synthesis of IMP in the breast muscle was different from thigh muscle maybe or that other ingredients of SBL played a role in part in the deposition of IMP. There are currently no data on the effect of SBL and the underlying mechanism of its action. In light of these results, future studies will evaluate the other mechanisms of SBL involved in improving meat flavor during regular temperature and heat stress.
In a summary, a basal diet supplemented with SBL increased muscle IMP content of AA broilers compared with the control, thereby, decreased the loss of meat flavor substance caused by heat stress. The likelihood was that SBL played a part in improving meat flavor during heat stress in AA broilers via enhancing the ADSL gene expression in livers.
ACKNOWLEDGEMENTS
The investigation was supported by the Special Fund for Technological Innovation Talents of Harbin Science and Technology Bureau (2011RFLXN015), the Program for Backbone Teachers Innovation Ability of Heilongjiang Province (1152G008), and the Program for Innovative Research Team of Northeast Agricultural University (IRTNEAU, CXT006-2-1).