Historical Overview of the Effect of β-Adrenergic Agonists on Beef Cattle Production
Article information
Abstract
Postnatal muscle hypertrophy of beef cattle is the result of enhanced myofibrillar protein synthesis and reduced protein turnover. Skeletal muscle hypertrophy has been studied in cattle fed β-adrenergic agonists (β-AA), which are receptor-mediated enhancers of protein synthesis and inhibitors of protein degradation. Feeding β-AA to beef cattle increases longissimus muscle cross-sectional area 6% to 40% compared to non-treated cattle. The β-AA have been reported to improve live animal performance, including average daily gain, feed efficiency, hot carcass weight, and dressing percentage. Treatment with β-AA increased mRNA concentration of the β2 or β1-adrenergic receptor and myosin heavy chain IIX in bovine skeletal muscle tissue. This review will examine the effects of skeletal muscle and adipose development with β-AA, and will interpret how the use of β-AA affects performance, body composition, and growth in beef cattle.
INTRODUCTION
Increasing lean yield, i.e., muscle mass, while also increasing feed efficiency would greatly benefit the beef cattle industry. One unique physiological characteristic of skeletal muscle tissue compared to other tissues is that the number of multinucleated cellular units, muscle fibers, is fixed at birth. This obviates hyperplastic growth as a means of increasing muscle mass, so increasing muscle hypotrophy has been a primary focus of research scientists at universities, government agencies, and pharmaceutical companies. Post-mitotic growth of muscle is achieved through either DNA accumulation or through increasing the ratio of protein to DNA in muscle fibers. Whereas steroidal implants tend to increase muscle mass by increasing the accumulation of DNA in muscle, beta-adrenergic agonists (β-AA) increase muscle mass by increasing the ratio of protein to DNA (Walker et al., 2010), similar to effects seen with the administration of somatotropin to beef cattle (Vann et al., 1998). Steroidal implants and β-adrenergic receptors (β-AR), commonly used in the North and South American cattle industries, have similar physiological responses of muscle hypertrophy, appear to have different mechanisms of action to achieve muscle growth, but the mode of action of β-AA in beef cattle is still not fully understood. Even less is known about the mechanisms by which β-AA depress adipose accretion in livestock species. This review will focus primarily on the biology of β-AA as it relates to lipid metabolism in adipose tissue and protein metabolism in skeletal muscle of livestock species.
PHYSIOLOGICAL FUNCTION OF β-ADRENERGIC AGONISTS
The α-adrenergic receptor (α-AR) and the β-AR are members of a large superfamily of G protein-coupled receptors (GPCR). Previous articles by Mersmann and coworkers have provided an overview of β-AR biology in livestock species, although these articles focused primarily on adipose tissue metabolism in swine (Mills and Mersmann, 1995; Mersmann, 1998; Mersmann and Smith, 2005). All β-AR contain seven hydrophobic, transmembrane domains (Figure 1). The β-AR are part of a large family of GPCR, which arose from a common ancestral gene, and were modified to accommodate a wide variety of ligands (Mills and Mersmann 1995). The homology among the receptors is approximately 50%, whereas across species, the homology is >75% within a subtype.

Model of the β2-adrenergic receptor.
In general, β-ARs consist of over 400 amino acids. Each β-AR contains three extracellular segments associated with the N-terminus and three intracellular loops with the C-terminus (Mills and Mersmann, 1995). Binding of the signal transduction elements occurs at intracellular loop 3 in the G-proteins, whereas the regulatory phosphorylation sites are located at the C-terminal. Phosphorylation at loop 3 leads in inactivation of the C. Ricks et al. (1984) proposed that β-AA reduce adipose tissue accretion by the combined effects of inhibiting de novo fatty acid biosynthesis and concurrently stimulating triacylglycerol (TAG) hydrolysis, or lipolysis (Figure 2). Conversely, muscle mass is increased by inhibiting protein turnover and promoting myofibrillar protein synthesis. The biological reactions of β-AA occur in peripheral tissues or cells that express β-AR. Most of the actions of β-AA can be attributed to an elevation of cAMP, as interaction of β-AA with β-AR stimulates adenylate cyclase via the stimulatory Gs-protein system (Figure 3). Once the concentration of cAMP is increased, it binds to the regulatory subunits of protein kinase A, which phosphorylates serine residues of several metabolic hormones (Mersmann, 1998). The phosphorylation of hormone sensitive lipase activates the enzyme, resulting in the partial hydrolysis of TAG, whereas the phosphorylation of acetyl-CoA carboxylase inhibits de novo biosynthesis of fatty acids. Thus, the overriding biological response of adipose tissue to β-AA is a depression of TAG accumulation and concomitant release of fatty acids (Oscar, 1995).
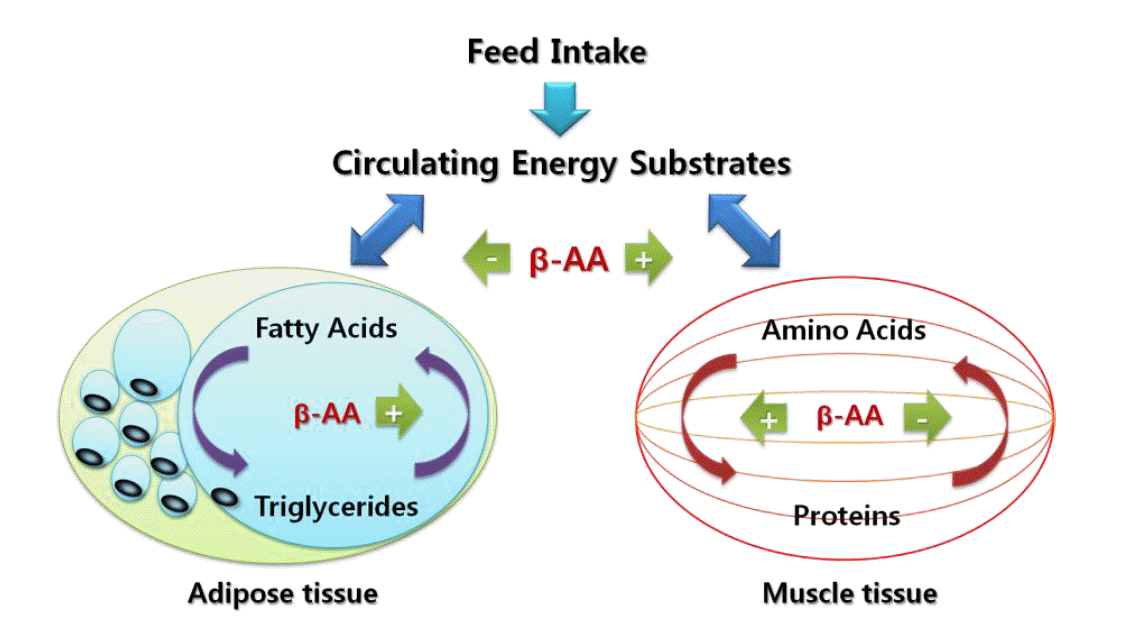
Proposed mechanisms of action by which β-adrenergic agonists decrease adiposity and increase muscle growth. Adapted from Ricks et al. (1984).
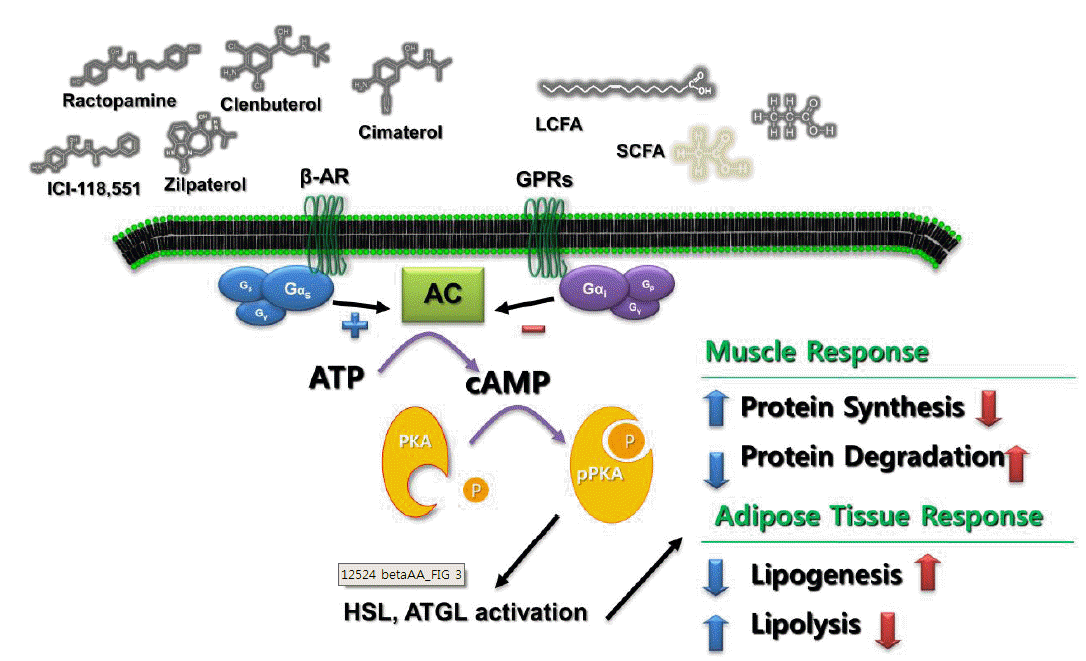
Interaction of proteins that mediate lipid mobilization from adipocytes. The first step in characterizing adrenergic agonists was to separate GPRs from β-ARs. While β-ARs regulate cell metabolism via Gs-protein stimulated cAMP-mediated events, GPRs oppose these effects because the GPR is coupled to the inhibitory Gi protein. Beta AA activated protein synthesis through the cAMP signal cascade in the skeletal muscle. AC, adenylate cyclase; ATGL, adipose triglyceride lipase; ATP, adenosine triphosphate ; cAMP, cyclic adenosine monophosphate; GPR, G protein-coupled receptor; HSL, hormone-sensitive lipase; PKA, protein kinase A; TG, triacylglyerol.
Historically, the first step in characterizing adrenergic agonists was to separate α-AR from β-AR (Sillence et al., 2005). Whereas β-AR regulate cell metabolism via Gs-protein stimulated cAMP-mediated events, α-AR oppose these effects because the α-AR is coupled to the inhibitory Gi protein (Figure 3). Epinephrine is the naturally occurring ligand for both the β-AR and α-AR, so the net response to epinephrine will depend on the relative abundance of the two receptors. However, the α-AR is not present in cell membranes of adipose tissue in livestock species, so this level of regulation is not essential to adipose tissue metabolism. After the initial demonstration of the β-AR, β1-AR subtypes were distinguished from β2-AR and β3-AR subtypes. It has also been suggested that a fourth β-AR may exist in porcine adipose tissue (Sillence et al., 2005). Granneman et al. (2001) suggested that the putative β4-AR could be a novel conformational state of the β1-AR. In addition, no unique sequence has been determined for the hypothesized fourth β-receptor. This suggests that it may be a novel state of another receptor subtype rather than a unique subtype. All three β-AR subtypes (β1, Accession #AF188187; β2, Accession #NM174231; β3, Accession #X85961) have been cloned and sequenced for bovine. A summary of the distribution of the β-AR subtypes in several species and tissues is listed in Table 1.

Distribution of β-AR subtypes
RESPONSIVENESS OF BOVINE TISSUES TO β-ADRENERGIC AGONISTS
The β-AR have been demonstrated in every cell type for every species investigated, but they differ widely in their distribution and specificity for synthetic ligands (Table 2). CGP20,712 is a β1-specific antagonist, whereas ICI118,551 is a specific antagonist for β2-AR. Notably, ICI118,551 has no effect in pigs. BRL37,344 is a β3-specific agonist in rodents, but is a β2-agonist in pigs. CGP12,177 stimulates basal lipolysis in rat adipocytes, as well as inhibiting response to isoproterenol.

Characterization of synthetic β-adrenergic agonists (AR) and antagonists in adipose tissues of rats, pigs, sheep, and cattle
Previous studies have investigated the effect of β2-AA on the individual muscle fiber level. Many of these studies assessed relative changes in both number as well as diameter of type I vs type II muscle fibers. Clenbuterol-fed heifers had greater type II muscle fiber diameters, but numerically type I muscle fiber diameters (Miller et al., 1988), relative to control heifers. Clenbuterol-fed steers also increased type IIA muscle fibers compared to control cattle (Smith et al., 1995). In addition, Kim et al. (1987) reported that in lambs fed cimaterol the proportion of type I to type II fibers in the longissimus and semitendinosus muscle were unaffected. However, the type II fibers had 50% greater cross-sectional area in the longissimus and semitendinosus muscle of cimaterol-fed lambs compared to untreated lambs. However, cimaterol had no effect on the cross-sectional area of type I fibers (Kim et al., 1987). These data from both cattle and sheep suggested that β2-AA can increase individual muscle fiber cross-sectional area. It is also apparent that type II fibers were the most responsive to the β2-AA administration. In another study Beermann et al. (1987) reported that dramatic increases in hind-limb muscle mass of lambs fed cimaterol was a result of radial growth of both type I and type II fibers. In both of these studies, authors found a reduction in proportion of type I fibers compared to type II fibers due to cimaterol administration. Due to these findings, the authors concluded that hypertrophy of type II fibers contributed more to the cimaterol-induced increase in skeletal muscle mass compared to type I hypertrophy. However, cimaterol administration resulted in preferential increase in the number of type II fibers compared to type I fibers. Zilpaterol hydrochloride (ZH), an orally active β-AA for finishing beef cattle, had no effect on β1-AR mRNA levels, but there was an increase in β2-AR mRNA levels (Baxa et al., 2010). Feeding of ZH the last 30 d prior to slaughter increased mRNA levels of myosin heavy chain-IIX (MHC-IIX) in semimembranosus muscle but had no detectable effect on MHC-I and IIA mRNA levels. These data indicate that the change of mRNA concentration of MHC isoform and β-AR are correlated with skeletal muscle hypertrophy of beef cattle.
Park et al. (2009) indicated that the phosphorylation and subsequent activation of AMPK (Adenosine Monophosphates-activated Protein Kinase), a protein kinase involved in energy metabolism, is positively correlated with increasing mRNA levels of MHC IIX in pigs. This data reported that injection of an AMPK activator (AICAR) increased phosphorylation of AMPK and synergistically enhanced mRNA of MHC IIX without affecting MHC I or IIA mRNA levels. Because AMPK phosphorylation can be regulated by cAMP (Park et al., 2009), this data supports the theory that the cellular mechanism of postnatal muscle hypertrophy can be influenced by β-AA.
THE β-ADRENERGIC AGONIST-STIMULATED MUSCLE HYPERTROPHY
Many β-AA can be fed to ruminants and the effects of feeding a β2-AA include the improvement of skeletal muscle mass and/or cross-sectional area of muscle fibers. Examples of these skeletal muscle-enhancing characteristics are summarized in Table 3.

Relative changes in individual muscle size and/or mass due to administration of a β2-adrenergic agonist to ruminants
Addition of either clenbuterol or cimaterol, both presumed β2-AA, to ruminants resulted in increased longissimus muscle (LM) cross-sectional area between 11% and 39%. In addition, the mass of selected muscles was increased between 8% and 40%. These data suggest that the addition of these β2-AA to ruminants results in a dramatic increase in muscle mass.
For β-AA to have direct effects on skeletal muscle hypertrophy, specific high-affinity receptors for the β-AA must be present on the surface of skeletal muscle cells. Previous research has reported the presence of both the mRNA for various subtypes of β-AR and specific binding sites for β-AA. Sillence et al. (1991) found that muscle homogenates from the beef LM contained predominately β2-AR. Specifically, the sarcolemma membrane preparations of the LM were shown to bind to the [125I] ICYP radio ligand, a radioactive ligand that binds β-AR with binding affinities consistent with β2-AR populations. In contrast, binding experiments revealed that there was no detectable β1-AR present in these sarcolemma membrane preparations (Sillence et al., 1991). Subsequent research by this group (Sillence and Matthews, 1994) confirmed that β1-AR could not be detected in bovine skeletal muscle and adipose tissue using classical ligand binding techniques. Other cell culture-related techniques have revealed that a very small population of β1-AR is detectable in bovine tissues and are coupled to the production of the second messenger, cAMP.
Research from our laboratory, utilizing quantitative real-time PCR, has detected mRNA for all three β-AR subtypes in total RNA isolated from bovine longissimus and semimembranosus biopsy samples (Baxa et al., 2010). It is noteworthy that on the average 96% to 99% of the β-AR mRNA was identified as β2 with the remaining 1% to 4% being β1 and β3 mRNA. The high abundance of β2-AR mRNA in bovine muscle tissue was consistent with ligand binding studies suggesting that greater than 90% of β-AR in bovine skeletal muscle.
THE β-ADRENERGIC RECEPTOR INCREASES DURING PHYSIOLOGICAL GROWTH
The presence of β-AR and distribution of different subtypes in skeletal muscle may be related to relative maturity of the animal. Minimal levels of β-AR were detected in multinucleated muscle cell cultures established from fetal bovine skeletal muscle as indicated by ligand binding studies. Additionally, short-term isoproterenol exposure did not increase cAMP production suggesting the lack of functional β-AR in fetal skeletal muscle (Bridge et al., 1998). However, small amounts of β2-AR mRNA were present in total RNA from these cultures (Bridge et al., 1998). Studies feeding clenbuterol to both young and old rats suggested that older, more mature rats were more responsive to clenbuterol administration compared to younger rats (Greife et al., 1989). Clenbuterol feeding for 14 d to older, heavier (185 g) rats increased growth rate 11% compared to only a 22% increase in lighter (100 g), younger rats (Greife et al., 1989). The authors speculated that the disparate response was related to relative differences in body fat content. However, it also is possible that the increased density of β-AR in older rats could have mediated the improvements in growth rate. Similarly, field reports suggest older, heavier yearling cattle are more responsive to a β-AA compared to younger, immature calf-fed cattle. However, Schiavetta et al. (1990) fed 0.5 ppm clenbuterol for 50 d to Angus steers beginning at 8 mo of age and demonstrated the most profound effects of β-AA reported for any study involving livestock species. In their study, clenbuterol increased feed efficiency by 25% and increased LM mass by 24%. Clenbuterol treatment also reduced marbling scores and carcass quality grades by one-half grade (from Choice0 to Select+) and increased cooked beef shear force by over 1 kg; the negative effects on carcass quality grade and beef tenderness persisted even after 78 d withdrawal from clenbuterol treatment (Schiavetta et al., 1990).
To further increase the understanding of the apparent age-dependent effects of β-AA administration on cattle growth, we determined the abundance of β-AR mRNA in muscle biopsies from calf-fed vs yearling-fed steers. Our preliminary data suggested that in yearling-fed steers, as days on feed increased, the abundance or β1-AR mRNA decreased and the levels of β2-AR mRNA significantly increased in skeletal muscle tissue (data not shown). In contrast, no change was detected in the mRNA levels of β1- or β2-AR in muscle biopsies from younger calf-fed steers. These data suggested that in older, heavier cattle, β2-AR mRNA increased with advancing days on feed thus altering functional β2-AR levels in skeletal muscle and ultimately may impact the responsiveness of skeletal muscle to β-AA.
In adipose tissue, isoproterenol (a selective β-AA agonist) has a strong lipolytic effect when infused through an i.v. in heifers (Ferlay and Chilliard, 1999) and lambs (Ferlay et al., 2001). Terbutaline, a β2-AA, has a slight lipolytic effect when infused in heifers and lambs, whereas dobutamine, a β1-AA, has no effect on lipolysis in heifers. The β3-AA CL316,243 has no effect on lipolysis in ewes (Ferlay et al., 2001), consistent with the lack of functional brown adipose tissue in postnatal lambs (Smith et al., 2004; Smith and Carstens, 2005).
PROTEIN SYNTHESIS IN SKELETAL MUSCLE
The protein synthesis of porcine skeletal muscle has been shown to be enhanced by the treatment of ractopamine (Bergen et al., 1989). It has also been reported that ractopamine treatment of a myotube culture (L6 cell culture model) increased myosin heavy-chain protein synthesis (Anderson et al., 1990). These data indicate that ractopamine induces muscle protein accretion and increased the myofibrillar and total protein synthesis. Treatment of ractopamine, a β1-AA, did not change protein degradation in cultured L6 muscle cells (Anderson et al., 1990). Thus, this manuscript concluded that ractopamine-enhanced muscle protein accretion was due to increased protein synthesis with no detectable effect on protein degradation. Additionally, both ractopamine and clenbuterol have been shown to increase myosin light chain-1 (the Type II MLC-1 isoform) mRNA abundance in the LM of beef cattle compared to untreated cattle (Smith et al., 1989; Smith et al., 1995). Increasing myofibrillar protein synthesis likely is a result of β-AA-induced up-regulation of myofibrillar protein gene transcription. The increased mRNA transcription of muscle protein indicated an elevation in muscle hypertrophy. McMillan et al. (1992) reported that clenbuterol stimulated protein synthesis in murine primary culture system. These conclusions are further supported by muscle cell culture studies in which β-AA stimulated protein synthesis.
PROTEIN TURNOVER IN SKELETAL MUSCLE
An increase in myofibrillar protein degradation can decrease net protein accretion in skeletal muscle if not compensated by increased protein synthesis. The rate of protein degradation has been determined by differences between protein accretion and fractional synthesis (Beermann, 2002). The activities of calcium-dependent and lysosomal proteases and their inhibitors also have been determined following β-AA administration. Early studies indicated that muscle protein degradation is reduced or unaffected by β-AA administration. The majority of cattle work in the area of protein degradation has been conducted with the β-AA, L-644,969. This β2-AA caused a 27% reduction in fractional protein degradation rate in steers compared to untreated controls (Wheeler and Koohmaraie, 1992). Additionally, the activity of a specific inhibitor to the calpains, calpastatin, was elevated in muscle samples of steers fed L-644,969 (Wheeler and Koohmaraie, 1992: Killefer and Koohmaraie, 1994). The treatment with β-AA has been reported to decrease overall meat tenderness in lambs (Hamby et al., 1986) and beef cattle (Miller et al., 1988; Schiavetta et al., 1990). The muscle proteases and inhibitors that regulate protein degradation that are affected in vivo often follow parallel responses in the postmortem meat. Schroeder et al. (2003) reported that 200 mg·hd−1·d−1 of ractopamine fed to steers and heifers had no effect on meat tenderness but 300 mg·hd−1·d−1 fed to steers increased shear force values compared to control samples. These data indicate that at higher doses ractopamine, a presumed β1-AA, can bind to β2-AR and elicit a biological response consistent with that type of receptor.
It is possible that IGF-I could be mediating the increases in skeletal muscle hypertrophy due to increased protein synthesis and decreased protein degradation with β-AA. In lambs, cimaterol feeding (10 ppm) has been shown to reduce circulating IGF-I levels by 46.5% at 42 d and 21.5% at 84 d compared to untreated lambs (Beermann et al., 1987). Similarly, in more recent studies by Walker et al. (2006), ractopamine feeding to Holstein steers attenuated the TBA/E2-induced increase in circulating IGF-I. Additionally, ractopamine feeding decreased LM IGF-I mRNA concentrations compared to concentrations observed before ractopamine feeding (Walker et al., 2006). Furthermore, IGF-I is known to stimulate the proliferative capacity of muscle satellite cells, which in turn supports postnatal muscle hypertrophy, although β-AA do not increase muscle DNA content. More research is needed to confirm the reduction in local and/or circulating IGF-I is related to the disconnect in satellite cell proliferation and fusion during β-AA stimulated skeletal muscle hypertrophy. The ability to circumvent these relationships could potentially sustain greater degrees of muscle hypertrophy following administration of β-AA.
RACTOPAMINE AND ZILPATEROL AS A SPECIFIC β-ADRENERGIC AGONISTS
The ZH has been used United States, South Africa, Mexico, and Canada for improving beef cattle growth and feed efficiency. The use of ZH in finishing diets improved average daily gain (ADG), weight gain, hot carcass weight (HCW), ribeye area, and dressing percentage (Avendano-Reyes et al., 2006; Baxa et al., 2010; Vasconcelos et al., 2008). Although feeding ZH has less effects in heifers, ZH used in last 20 to 40 d before slaughter causes increased ADG, body weight (BW), gain to feed ratio (G:F), and HCW in steers and heifers (Montgomery et al., 2009).
Zilpaterol (Figure 4) is strikingly different in terms of its structure compared to other β2-AA like ractopamine, clenbuterol or cimaterol. The structures of clenbuterol and cimaterol are very similar to norepinephrine and epinephrine. Recent results indicated that ZH binds with high affinity to β2-AR, but the binding affinity of ZH to β2-AR was lower as compared to clenbuterol (10−6 M vs 10−8 M). Additionally, the binding of ZH to β1-AR was weaker than its binding affinity to β2-AR (Verhoeckx et al., 2005). Subsequent work has confirmed these results and extended their understanding by showing that ZH, acting through a β2-AR, increased cAMP levels in cultured U937 macrophages (Verhoeckx et al., 2006).
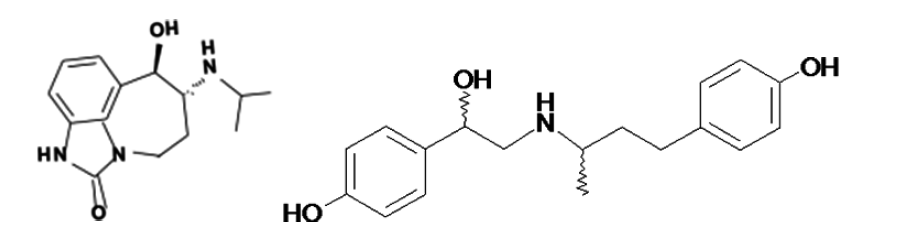
Structure of Zilpaterol (RU 42173) and Ractopamine.
PRACTICAL CONSIDERATIONS
The maximal response of β-AA is affected by the dose and duration of a sustained dose (e.g., in its effects on MLC-1 gene expression; Smith et al., 1989). It has been well documented that the β-AR-mediated increase in cAMP is transient and that continuous receptor activation by the β-AA ligand is necessary to maintain the cAMP levels to sustain the response. Exposure to a constant dose of β-AA to the receptor will eventually cause acute desensitization or inactivation of receptor-mediated signaling. The phosphorylation of both protein kinase A and β-AR kinases are the main steps in initiating the signaling cascade following β-AR activation of cAMP (Hausdorff et al., 1990). Acute desensitization can be circumvented to some degree by increasing the dose and potentiating the signal. Long-term exposure (chronic) at elevated doses of β-AA leads to internalization or loss of the receptor from the cell surface and down-regulation in β-AR mRNA abundance (Hausdorff et al., 1990). These alterations appear to be irreversible at least in the short-term feeding period.
SAFETY CONSIDERATIONS WITH UNAPPROVED-β-ADRENERGIC AGONISTS
Both clenbuterol and salbutamol are presumed β2-AA that have been studied as growth promotants in many species including beef cattle. The biological effects of clenbuterol in beef cattle were first described by Ricks et al. (1984). Clenbuterol is chemically described as a benzyl alcohol, 4 amino-alpha-(tbutylamino)methyl-3,5-dichloro. Earlier research studies have indicated that clenbuterol was a very potent β2-AA in cattle. Very little information is available in the scientific literature in regards to salbutamol.
Neither clenbuterol nor salbutamol is approved for use in meat animal production in the USA. The pharmaceutical company, American Cyanamid, abandoned the approval process for clenbuterol in the early 1990s when it appeared there may be a residue toxicity issue with clenbuterol in certain edible tissues of beef. Clenbuterol received much negative attention in the early 1990s when its illegal feeding to cattle resulted to cases of food poisoning in both Spain and France (Martinez-Navarro, 1990; Salleras et al., 1995). Individuals consuming liver from animals illegally-fed clenbuterol possessed symptoms of tachycardia, tremors, headache, and dizziness.
Indiscriminate use of clenbuterol and other non-approved β2-AA such as salbutamol continue to be a major issue across the globe. Individuals who illegally feed these agents to cattle often do so at rates much greater than were ever tested in research trials. This can result in residue levels in both the kidneys and liver at levels great enough to cause food poisoning. Research by Smith and Paulson (1997) revealed that clenbuterol residues were greatest in lung, kidney, and liver in cattle fed 3 mg/kg (3 ppm) BW of clenbuterol. These concentrations averaged 8.36 ppm in lung tissue, 5.9 ppm in kidney, and 5.04 ppm in liver. It has been suggested that the effective oral dose of clenbuterol in humans can be as low as 10 μg/d. Levels above this could begin to cause food poisonings. At a residue of 5 ppm, an individual would only need to consume 2 g to equal the effective dose in humans. Due to these human health concerns, clenbuterol and other potent β2-AA will most likely never be approved for use in meat-animal production.
The two β-AA that have been approved for use in the U. S. and other countries around the globe, ractopamine and ZH, have been studied extensively from a residue standpoint. When fed at the approved dose and label directions are followed for withdrawal times, negligible to undetectable levels of residues have been reported. For example, ZH feeding at the normal approved dose resulted in liver and kidney residue levels of 0.18 and 0.09 ppm after a 48 h withdrawal (Zilmax FOI Summary, NADA 141–258, August, 2006). These absolute concentrations are much less than those reported for clenbuterol. Additionally, the effective oral dose of ZH in humans is much higher than that reported for clenbuterol. Taken together, these data indicate that there are key differences in the metabolism, excretion and activity of different β-AA.
CONCLUSIONS
As indicated in this review, β-AA consistently improve beef cattle performance and increase muscle growth when mixed with finishing rations. The β-AA increase ADG, G:F, HCW, LM cross-sectional area and mass, and dressing percentage. However, cattle receiving β-AA tend to have a reduced marbling scores, lower back-fat thickness and increased beef toughness. Elevation of glycolytic fiber types with β-AA treatment is primarily responsible for the increased muscle hypertrophy but otherwise is negatively correlated with both intermuscular and intramuscular adipose tissue amount. Further study is needed to find methods to enhance growth performance without reducing beef quality in β-AA-treated cattle.
ACKNOWLEDGMENTS
This work was carried out with the support of “Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ009414 )” Rural Development Administration, Republic of Korea.