Dragon fruit (Hylocereus undatus) peel pellet as a rumen enhancer in Holstein crossbred bulls
Article information

Abstract
Objective
An experiment was conducted to assess the effect of dragon fruit peel pellet (DFPP) as a rumen enhancer of dry matter consumption, nutrient digestibilities, ruminal ecology, microbial protein synthesis and rumimal methane production in Holstein crossbred bulls.
Methods
Four animals, with an average live-weight of 200±20 kg were randomly assigned in a 4×4 Latin square design to investigate the influence of DFPP supplementation. There were four different dietary treatments: without DFPP, and with 200, 300, and 400 g/h/d, respectively.
Results
Results revealed that dry matter consumption of total intake, rice straw and concentrate were not significantly different among treatments (p>0.05). It was also found that ruminal pH was not different among treatments (p>0.05), whilst protozoal group was reduced when DFPP increased (p<0.01). Blood urea nitrogen and NH3-N concentrations were increased at 400 g of DFPP supplementation (p<0.01). Additionally, volatile fatty acid production of propionate was significantly enhanced by the DFPP supplementation (p<0.05), while production of methane was consequently decreased (p<0.05). Furthermore, microbial protein synthesis and urinary purine derivatives were remarkably increased especially at 400 g of DFPP supplementation (p<0.05).
Conclusion
Plant secondary compounds or phytonutrients (PTN) containing saponins (SP) and condensed tannins (CT) have been reported to influence rumen fermentation. DFPP contains both CT and SP as a PTN. The addition of 400 g of DFPP resulted in improved rumen fermentation end-products especially propionate (C3) and microbial protein synthesis. Therefore, DFPP is a promising rumen enhancer and indicated a significant potential of DFPP as feedstuff for ruminant feed to mitigate rumen methane production.
INTRODUCTION
Livestock production contributes to global warming by discharging greenhouse gas (GHG) either from manure management and enteric fermentation or feed production. The major GHGs are nitrous oxide (N2O), carbon dioxide (CO2) and especially methane (CH4) throughout the production process. Therefore, CH4 is the most important GHGs from the livestock [1].
The importance of ruminal ecology has been stated to the subsequent fermentation and livestock production efficiency [2]. Manipulation of ruminal fermentation has been reported using ionophores, antibiotics, chemical compounds, vaccination, and phytonutrients or plant secondary compounds [3]. Moreover, there is a need to utilize organic plant products that are used as livestock feed additives to improve the efficiency of feed use in ruminant production, and to decrease methane emission [4]. Natural products in tropical regions contain high to medium amounts of phytonutrients especially condensed tannins (CT) and saponins (SP), which have activity against protozoal populations and methane emission [5]. Previous studies have stated that the use of agricultural by-products and tropical plants especially fruit peel wastes containing phytonutrients have resulted in improved ruminal fermentation while decreasing production of methane [6,7]. Currently, plant secondary substances are an important area of research to replace feed additives in livestock [8]. The CT and SP are a group of phytonutrients of that demonstrate an ability to alter the rumen ecosystem, fermentation, reduces production of methane, and increases livestock production [9].
Among many tropical fruits, dragon fruit is abundant and its fruit peel is a waste from the fruit industry or fruit processing and is traditionally used as fertilizer. The peel has high amounts of phytonutrient [10]. Hence, dragon fruit can be considered as a good source of bioactive compounds for fermentation modification and health benefits. In addition, CT and SP are phytonutrients contained in dragon fruit peel and in many tropical plants [11]. In addition, Matra et al [12] studied in vitro fermentation by adding at 0%, 1%, 2%, 3%, and 4% dragon fruit peel powder containing 6.9% CT and 8.9% SP, resulting in improving ruminal fermentation and decreasing protozoal population and methane production with the 4% dragon fruit peel powder addition. However, there are limited studies on the impact of dragon fruit peel pellet (DFPP) containing CT and SP as a supplement on nutrient digestibility, rumen fermentation, and methane emission. Therefore, the study objective was to explore the effect of DFPP as a rumen enhancer on dry matter (DM) consumption, nutrient digestibilities, ruminal ecology, microbial protein synthesis and methane emission in Holstein crossbred bulls.
MATERIALS AND METHODS
Animal care
All Holstein crossbred bulls involved are managed and cared for according to the protocols of the Animal Care and Use Committee of Khon Kaen University and carried out by the Institute of Animals for Scientific Purpose Development (IAD), Thailand (record no. U1-06878-2560).
Animal management
The experimental animals were injected with vitamin AD3E and drenched with Ivermectin (1 mL/50 kg live-weight [LW]; Merck Co., Inc., Kenilworth, NJ, USA) before imposing the respective treatments. They were housed in individual pens with clean drinking water and mineral blocks were made available freely.
Preparing of dragon fruit peel pellet
Fresh dragon fruit peels were obtained from fruit canning factory. They were chopped and sun-dried approximately 2 or 3 days. Dried dragon fruit peels were processed by grinding to the length of 1 mm sieve (Cyclotech Mill, Tecator, Höganäs, Sweden). The powder was mixed with other components (Table 1) to form a pellet by using Ryuzo-kun pelleter (Kakiuchi Co., Ltd., Nankoku, Kochi, Japan).
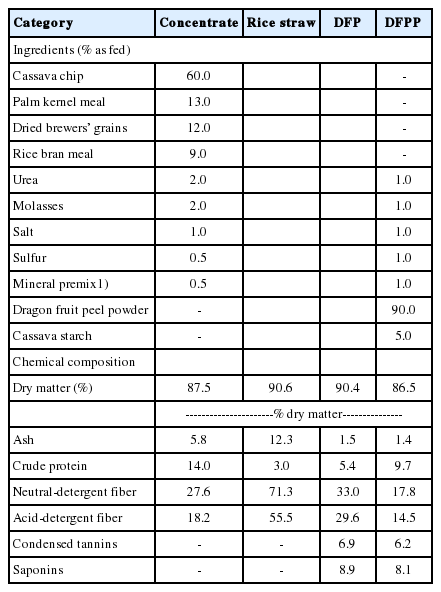
Ingredients and chemical composition of the experimental feedstuffs
Experimental design and dietary treatments
Four Holstein crossbred bulls with an average LW of 200±20 kg were used. The experimental design was a 4×4 Latin square design with four different treatments (0, 200, 300, and 400 g/h/d DFPP, respectively). Animals were subjected to their dietary treatments during four periods consisting of 21 days per period; the first 14 days were used for diet adaptation and the last 7 days were used for data collection. The last 7 days, animals were moved to metabolism crates for total fecal and urine collection to study digestibility. Animals were offered rice straw as a roughage source ad libitum with 10% refusal and 0.5% concentrate of their body weight comprising 14.0% crude protein (CP), 27.6% neutral detergent fiber (NDF), and 18.2% acid detergent fiber (ADF) at 07.00 and 16.00 o’clock.
Sample collection, preparation and chemical analyses
Urinary and fecal samples were collected during the last 7 days of each period when the animals were in metabolism crates. The feces were dried at 60°C for 2 or 3 days then were ground (Cyclotech Mill, Tecator, Höganäs, Sweden) and used for nutrient digestibilities. The concentrate, rice straw, dragon fruit peel and feces were chemically assessed for DM, ash, CP, and fiber contents. The DFPP samples were measured for CT and SP content following the procedure of Wanapat et al [13].
The urinary allantoin was analyzed by the procedure of Chen et al [14]. Microbial purine absorbed corresponding to the purine derivatives (PD) excretion was evaluated based on their relationship according to Chen and Gomes [15] as follows:
where Y is the urinary PD excretion (mmol/d); X is the microbial purine absorbed (mmol/d); and W is BW of animal (g/kg BW0.75).
The microbial protein supply in grams per day were cal culated following to the equation:
where X is the urinary PD absorption (mmol/d), according the suppositions calculated by Chen and Gomes [15]. The microbial purine digestibility is 0.83 and the N concentration of purine is 70 mg N/mmol. The ratio of purine-N to total N in mixed rumen microbes is 11.6:100.
Efficiency of microbial N synthesis (EMNS) was calculat ed according to the following formula:
where DOMR is rumen digestible organic matter (OM) apparently fermented (assuming that rumen digestion was 650 g/kg OM of digestion in total tract, DOMR = DOMI×0.65, DOMI = digestible OM intake according to the method of ARC [16].
On the last day of each period, at 0 h before feeding and 4 h after feeding, rumen fluid was taken via a tube connected to a vacuum pump. Firstly, the rumen fluid samples were immediately determined for temperature and pH (HANNA Instruments HI 8424 microcomputer, Singapore). Secondly, rumen fluid of 1-mL was mixed with 10% formalin of 9-mL for measuring total direct counts of bacteria, protozoa and fungi zoospore according to the procedure of Galyean [17]. Thirdly, 45-mL rumen fluid were mixed with 1 M sulphuric acid of 5-mL to prevent the fermentation process and kept in plastic bottle, and then centrifuged at 3,000×g for 10 minutes. After the centrifugation, the samples were used for NH3-N analysis (Kjeltech Auto 1030 analyzer, Tecator, Hoganiis, Sweden) following the method of AOAC [18] and volatile fatty acid (VFA) analysis using high performance liquid chromatography (Model Water 600; UV detector, Millipore Crop, Cortland, NY, USA) according to Samuel et al [19] procedure. The methane production was estimated from VFA profiles following the method of Moss et al [20].
Blood samples at 0 and 4 h after feeding were collected (10 mL from the jugular vein) on the last days and then added with ethylenediaminetetraacetic acid. They were centrifuged for 20 min to separate plasma and stored at −20°C for blood urea nitrogen (BUN) analysis [21].
Statistical analysis
All data were analyzed according to the procedure of general linear model following to SAS [22]. The results are presented as mean values with the standard error of the means. Differences among treatment means with p<0.05 and p<0.01 were reported as being statistically different. All treatment means were analyzed according to the equation:
where μ, over all sample mean; Ti, effect of treatment i; Cj, effect of treatment i at column j; Rk, effect of treatment i at row k; eijk, error; and Yijk, the criteria under study, in treatment i, column j, row k.
Treatment means were determined by Duncan’s new mul tiple range test according the standard method of Steel and Torrie [23]. Additionally, orthogonal polynomials for diet responses were detected by linear, quadratic and cubic effects.
RESULTS
Chemical composition of experimental diets
Nutritive values of feeds are presented in Table 1. Concentrate was formulated to contain 14.0% CP, 27.6% NDF, and 18.2% ADF. Cassava chip (60% as fed) was used as an energy source. While, rice straw (3.05 CP, 71.3% NDF, and 55.5% ADF) was fed as a roughage source. Additionally, DFPP has 9.7% CP, 17.8% NDF, 14.5% ADF, with 6.2% CT and 8.1% SP, respectively.
Dry matter consumption and nutrient digestibilities
Table 2 illustrates the information on DM consumption and nutrients digestibilities as affected by the DFPP supplementation. The DM consumption of total intake, rice straw and concentrate were not affected by the DFPP supplementations (p>0.05). Moreover, nutrient digestibilities (DM, OM, CP, NDF, and ADF) were not affected (p>0.05) by the DFPP supplementation.
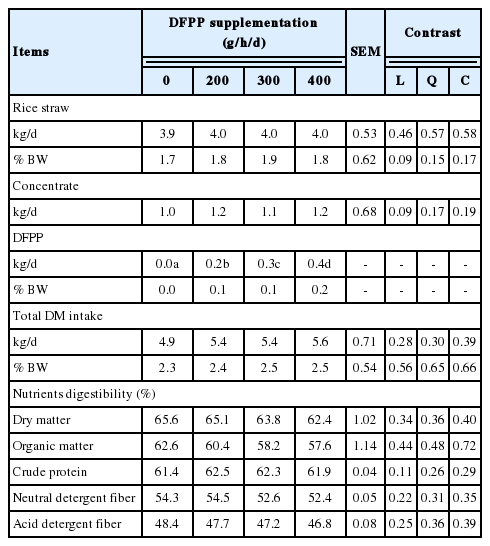
Effect of dragon fruit peel pellet supplementation on dry matter consumption and nutrient digestibilities in Holstein crossbred bulls
Ruminal ecology and blood metabolites
The effect of DFPP supplementation on ruminal ecology and BUN concentration in Holstein crossbreed bulls is shown in Table 3. The ruminal temperature and pH were not altered by the DFPP supplementation (p>0.05). The protozoa numbers at 0 h before feeding and 4 h after feeding were linearly (p<0.01) decreased by 69.5% and 65.7% and bacterial populations were enhanced when DFPP supplementation were increased up to 400 g (p<0.01). The DFPP supplementation levels did not affect the population of fungi (p>0.05). The DFPP supplementation significantly increased (p<0.01) the concentrations of NH3-N. Also, BUN at 0 h before feeding and 4 h after feeding were significantly increased (p< 0.01) by increasing DFPP supplementations (23.4% and 23.8%).
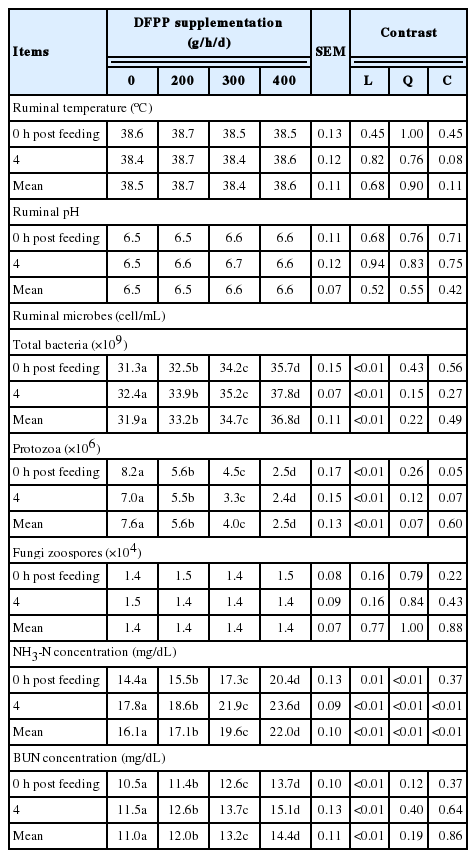
Effect of dragon fruit peel pellet supplementation on rumen characteristics and blood metabolites in Holstein crossbred bulls
Ruminal volatile fatty acid and methane production
The propionate concentrations at 0 h before feeding and 4 h post feeding were markedly increased (p<0.05) by the DFPP supplementation at 400 g/d when compared to the control group (11.8% and 12.2%). The concentration of acetate, butyrate and acetate to propionate ratio were significantly decreased by the DFPP supplementation (p<0.05). Total VFA were not influenced by the DFPP supplementation (p>0.05). The DFPP supplementations did not affect the methane concentration compared to the control, and DFPP supplementation at 400 g/d showed the lowest CH4 production of 9.0% at 0 h before feeding and of 10.0% at 4 h post feeding (Table 4).

Effect of dragon fruit peel pellet supplementation on ruminal volatile fatty acids production and methane production in Holstein crossbred bulls
Urinary purine derivatives and estimated microbial protein synthesis
Urinary PD, allantoin absorption and excretion were affected by the DFPP supplementation (p<0.05). Moreover, EMNS and microbial protein supply were improved by increasing level of DFPP supplementation especially at 400 g (p<0.05; Table 5).
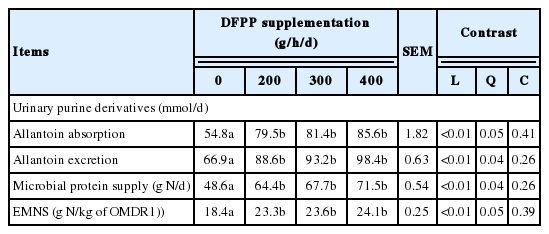
Effect of dragon fruit peel pellet supplementation on urinary purine derivatives and microbial protein synthesis in Holstein crossbred bulls
DISCUSSION
Dry matter consumption and nutrient digestibilities
The DM consumption was not affected by the DFPP supplementations. This could be due to the DFPP having no negative effect on the palatability of the diet. Ampapon et al [24] showed that DM consumption was not significantly different among groups of swamp buffalos fed different levels of mangosteen peel powder. Addition of plants containing phytonutrients into animal diets normally decreases digestion rate. It has been shown that nutrients’ digestibility were reduced by high intake of CT and SP level [25]. In this experiment, nutrients’ digestibility among treatments were similar. This could be due to the appropriate dose of CT and SP in DFPP, hence DFPP supplementation could be an alternative feed without a negative effect on nutrients’ digestibility. Similarly, Ampapon et al [26] revealed that the nutrients’ digestibility was not changed in the rambutan peel additional groups.
Ruminal ecology and blood metabolites
Russell and Rychlik [2] reiterated the important role of pH in maintaining normal activities in the rumen environment. Under this study, ruminal pH was 6.50 to 6.63, these values were not changed by adding DFPP at all levels. Accordingly, ruminal pH should be at 6.2 to 7.2 for microbial activity [27]. The protozoal population were decreased when adding DFPP, which could be attributed by the presence of phytonutrients especially CT and SP contained in the DFPP. These compounds could damage some enzymes and proteins on the plasma membrane of protozoa, causing the protozoa to die [28]. This finding agrees with Patra and Saxena [29], who reported that the population of protozoa was decreased by plant extracts (CT and SP) in the diets. Additionally, Matra et al [12] also explained that the inclusion of dragon fruit peel powder in the feed decreased protozoal population in in vitro study. Ruminal NH3-N concentrations were increased from 16.1 to 22.0 mg/dL by the DFPP supplementation. This may be due to the greater protein digestion from DFPP (9.7% CP), concentrate (14.0% CP) and non-protein nitrogen in the rumen [30].
Blood urea concentrations correlated with the ammonia nitrogen concentration availability in the rumen. Byers and Moxon [31], demonstrated that the optimal range of BUN was from 10.0 to 15.0 mg/dL. Under this study, BUN concentrations were increased when the cattle were fed with 400 g/h/d of DFPP, which ranged from 11.0 to 14.4 mg/dL. The reason of increasing BUN concentration could be due to the increase of ammonia nitrogen concentration. Accordingly, Foiklang et al [32] revealed that BUN concentration was higher in the mangosteen peel powder and grape pomace powder supplemented groups.
Ruminal volatile fatty acids and methane production
Under this experiment, adding levels of DFPP increased propionate whilst acetate, butyrate and acetate to propionate ratio were remarkably decreased. This may be attributed to when acetate reduced, propionate could be increased while propionate remained unaltered [33]. However, acetate inhibitory effect might decrease the acetate to propionate ratio and directed to inhibit ruminal methanogen [34]. Anantasook et al [35] stated that decreasing ratio of acetate to propionate related to the increase of propionate production from a biological conversion of hydrogen as a substrate. Furthermore, the ruminal VFA production differences may occur due to reduced fermentation rates resulting from the amount of tannins and/or SPs [36]. This finding aligns with the report of Piñeiro-Vázquez et al [37], who revealed that the propionate proportion was improved by 3% and 4% tannins extracted from Quebracho plant. Foiklang et al [30], who also found that the high propionate proportion was caused by reducing methane production due to the CT containing in grape pomace powder (12.3% CT).
The current study has shown that increasing the DFPP linearly reduced methane production (0 and 4 h post feeding). A reduction of methane from DFPP feeding could be due to the effect of the CT and SP. Accordingly, Hristov et al [38] and Gemeda and Hassen [39] also explained that the use of phytonutrient plants containing CT and SP had decreased the production of methane, decreased protozoal population and modified VFA in the rumen. Four possible functions of tannin on CH4 reduction have been proposed: i) it acts to reduce methanogen bacteria; ii) it decreases the protozoal population; iii) it acts to reduce fiber degradation; and iv) it decreases the acetate to propionate ratio [28]. Yang et al [40] showed that addition of tannic acid in beef cattle significantly reduced CH4 production.
Urinary purine derivatives and microbial protein synthesis
Urinary excretion is regarded as a marker of microbial protein synthesis in the rumen [41]. PD excretion and absorption were different among levels of DFPP supplementation, especially DFPP at 400 g had the highest value. Microbial protein supply under this trial, exhibited a quadratic increase. This could possibly be due to microbial protein supply and EMNS which may be reported that ruminal NH3-N produced was used for microbial activity and growth [42]. Firkins et al [43] explained that the synthesis of rumen microbial proteins would result in more proteins supplied to the small intestine of ruminants.
CONCLUSION
Supplementation of DFPP at 400 g/h/d resulted in an improvement of ruminal ecology, increasing VFA production especially propionate, microbial protein synthesis and reduction rumen methane production. Hence, DFPP could be a promising rumen enhancer and mitigate rumen methane production.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.
ACKNOWLEDGMENTS
The authors wish to sincerely express gratitude to Thailand Research Fund (TRF) through the Thailand Science Research and Innovation (TSRI) (TRF-IRN57W0002 and TRF-IRG5980010) and the National Research Council of Thailand (NRCT) through the Basic Research Fund (record no. 2564A10302002). Sincere thanks to Tropical Feed Resources Research and Development Center (TROFREC), Department of Animal Science, Faculty of Agriculture, Khon Kaen University, Thailand for the use of experiment facilities.