Effects of purified lignin on in vitro rumen metabolism and growth performance of feedlot cattle
Article information
Abstract
Objective
The objectives were to assess the effects of purified lignin from wheat straw (sodium hydroxide dehydrated lignin; SHDL) on in vitro ruminal fermentation and on the growth performance of feedlot cattle.
Methods
In vitro experiments were conducted by incubating a timothy-alfalfa (50:50) forage mixture (48 h) and barley grain (24 h) with 0, 0.25, 0.5, 1.0, and 2.0 mg/mL of rumen fluid (equivalent to 0, 2, 4, 8, and 16 g SHDL/kg diet). Productions of CH4 and total gas, volatile fatty acids, ammonia, dry matter (DM) disappearance (DMD) and digestion of neutral detergent fiber (NDF) or starch were measured. Sixty Hereford-Angus cross weaned steer calves were individually fed a typical barley silage-barley grain based total mixed ration and supplemented with SHDL at 0, 4, 8, and 16 g/kg DM for 70 (growing), 28 (transition), and 121 d (finishing) period. Cattle were slaughtered at the end of the experiment and carcass traits were assessed.
Results
With forage, SHDL linearly (p<0.001) reduced 48-h in vitro DMD from 54.9% to 39.2%, NDF disappearance from 34.1% to 18.6% and the acetate: propionate ratio from 2.56 to 2.41, but linearly (p<0.001) increased CH4 production from 9.5 to 12.4 mL/100 mg DMD. With barley grain, SHDL linearly increased (p<0.001) 24-h DMD from74.6% to 84.5%, but linearly (p<0.001) reduced CH4 production from 5.6 to 4.2 mL/100 mg DMD and NH3 accumulation from 9.15 to 4.49 μmol/mL. Supplementation of SHDL did not affect growth, but tended (p = 0.10) to linearly reduce feed intake, and quadratically increased (p = 0.059) feed efficiency during the finishing period. Addition of SHDL also tended (p = 0.098) to linearly increase the saleable meat yield of the carcass from 52.5% to 55.7%.
Conclusion
Purified lignin used as feed additive has potential to improve feed efficiency for finishing feedlot cattle and carcass quality.
INTRODUCTION
Inclusion of antibiotics in animal diets at sub-therapeutic levels can prevent disease and increase growth and improve feed efficiency [1]. However, such practices can select for antibiotic resistant bacteria and increase their concentrations in feces [2]. Consequently, there is a growing global response in livestock industries to restrict the sub-therapeutic use of antibiotics in livestock diets, especially those used to promote growth. Natural plant compounds may serve as alternatives to antibiotics, and although the impact of some of these has been assessed in vitro [3,4], none have been shown to consistently promote growth in vivo. Examples of natural plant compounds include tannins, saponins, and essential oils [5,6]. Lignin comprises a group of phenolic polymers that are essential components of plant cell wall structure. Globally, approximately 50 to 100 million tonnes of lignin are separated from wood during pulp and paper-making each year. However, only a small portion (1% to 2%) of this material is processed into value-added products with the majority being incinerated [7,8]. With the emergence of the cellulosic bio-ethanol industry, the global supply of lignin may further increase. It has been estimated that the global lignin market could reach 985.5 million US dollars by 2023 [9]. Although some purified lignins have antioxidant and antimicrobial activities [10–12], few studies have been conducted to evaluate their effects on nutrient digestion or growth in livestock. Phillip et al [13] observed improved growth in Holstein calves supplemented with organosolv lignin (Alcell lignin) at 12.5, but not at 25 or 50 g/kg diet dry matter (DM). Our previous work showed that a purified lignin from wheat straw (sodium hydroxide dehydrated lignin; SHDL) fed to lambs at 15 g/kg DM increased feed efficiency by 9% [14]. However, there is no information of SHDL on the growth performance of feedlot cattle.
The objectives of this study were to assess the effects of SHDL on in vitro ruminal fermentation of different type of diets and to evaluate the effects of SHDL as feed additive on the growth performance and carcass traits of feedlot cattle.
MATERIALS AND METHODS
Generation of sodium hydroxide dehydrated lignin
Sodium hydroxide dehydrated lignin was produced by extraction of wheat straw with aqueous sodium hydroxide at 160°C, and the resulting alkaline extract was acidified with mineral acid, filtered, washed with water and dried with the procedures described by Lora [15].
Determination of effects of sodium hydroxide dehydrated lignin on ruminal fermentation in vitro
Treatment, substrate and inoculum
Batch culture incubations were conducted to assess the effects of SHDL on ruminal fermentation of a mixed forage diet of ground (1.00 mm) alfalfa and timothy hay in the ratio of 50:50 (DM basis), and ground barley grain that passed a 2.0 mm sieve but was retained on a 0.5 mm sieve [16]. The SHDL was added to the forage and barley grain incubations at 0, 0.25, 0.5, 1.0, and 2.0 mg/mL (roughly equivalent to 0, 2, 4, 8, and 16 g/kg diet). Dosages of SHDL used in this experiment were based on previous study that showed SHDL supplemented at 15 g/kg diet improved feed efficiency for feedlot lambs [14].
The incubations were conducted independently for each substrate in 135-mL serum vials containing 500 mg of substrate and 50 mL of mixed rumen inoculum. The SHDL was added as a freshly prepared 1.0 mL suspension in distilled water (dH2O) at the concentrations of 12.5, 25, 50, and 100 mg/mL to achieve the respective dosages in the final incubation with mixed rumen microbial inoculum. Distilled water (1.0 mL) only was added to the control vials.
Rumen fluid was obtained from three rumen-cannulated, dry Angus cows fed (DM basis) a mixed ration of 65% of alfalfa-grass hay and 35% of barley grain along with 5% of a vitamin-mineral premix. For barley grain substrate, rumen fluid was collected from the same cows, but after they were adapted to the same diets as used in the finishing feedlot experiment. Rumen fluid and contents were collected from the dorsal, caudal, ventral and cranial regions of each animal 1 h before feeding, combined and strained through four layers of cheesecloth. Rumen fluid was combined across all cows and mixed with a mineral buffer [17] at a ratio of 1:2 as described by Wang et al [16]. All cows used in this study were cared for in accordance with standards of the Canadian Council on Animal Care [18].
Incubation and measurements
At the beginning of each incubation, inoculum was dispensed anaerobically (50 mL) into vials with substrate and immediately sealed and affixed to a rotary shaker (120 rpm) in an incubator at 39°C. Vials containing inoculum only were included as a negative control (blanks). Triplicate vials were prepared for each treatment with the entire experiment being repeated three times in separate weeks. Incubations were 48 h for forage and 24 h for barley grain. The volume of gas produced (GP) in each vial was measured after 3, 6, 9, 12, 24 (forage and barley grain), and 48 h (forage only), using a water displacement apparatus constructed in our laboratory, similar to a design described by Fedorak and Hrudey [19]. Gas samples (15 mL) were taken prior to release of headspace gas after each measurement and stored in an evacuated 6.8-mL container (Labco Ltd., High Wycombe, Bucks, UK) for later analysis of methane. At the end of incubation, fermentation residues were isolated by centrifugation (500×g, 10 min, 4°C) and washed three times by re-suspension in dH2O and collection by centrifugation [20]. Pellets were freeze-dried for determination of DM disappearance (DMD) and analysis of neutral detergent fibre (NDF) to estimate the 48-h in vitro NDF disappearance (NDFD; forage only). The liquid fraction was recovered by centrifugation (12,000×g, 15 min, 4°C) and the supernatant was used to determine NH3-N and volatile fatty acids (VFA).
Barley grain was processed in three different manners
i) particle size of 0.5 to 2.0 mm, ii) steam-rolled, and iii) kernels sectioned longitudinally. Each type of grain was incubated with SHDL at 0, 0.5, 1.0, and 2.0 mg/mL (roughly equivalent to 0, 4, 8, and 16 mg/kg diet DM). The treatments were arranged as 3×4 factorial design.
The incubation was conducted using the same procedure as described for mixed forage study, but the incubation was only conducted for 12 h because of the rapid degradation rate of the barley starch in the rumen. Gas production and in vitro DMD were also determined as described above. The incubation was repeated in three runs of incubations with two vials per treatment in each run.
Determination of effect of sodium hydroxide dehydrated lignin on animal performance of feedlot cattle
A total of 60 Hereford-Angus cross weaned steer calves (about 8 months old) were used in a feedlot experiment to evaluate the effects of SHDL on growth performance and carcass traits of feedlot cattle. Cattle (initial body weight 322.2±2.95 kg) were purchased from a local auction market and upon arrival at the Lethbridge Research and Development Centre, were ear tagged, branded, dewormed (Dectomax [doramectin, 0.5%], Pfizer Animal Health, Exton, PA, USA) and vaccinated against Infectious Bovine Rhinotracheitis (IBR), Parainfluenza-3 (PI3), Haemophilus somnus (Resvac 2/Somubac, Pfizer Animal Health, USA) and Clostridium spp. (Tasvax 8, Schering-Plough Animal Health, Upper Hutt, NZ). Steers were fed a barley silage-barley grain based total mixed ration (TMR) formulated to meet or exceed the nutrient requirements of beef cattle [21]. Steers were randomly assigned to four TMR diets (15 steers per diet) containing SHDL (GreenValue SA, Alpnach Dorf, Switzerland) at 0, 4, 8, and 16 g/kg diet DM (Table 4) and housed in individual pens.
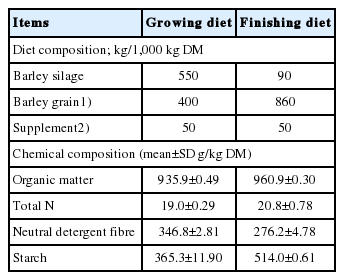
Composition of total mixed rations fed to steers in feedlot experiment
The experiment comprised a 70-d growing period, a 4-wk transition period in which the level of barley grain in the diet was increased and silage decreased at 7-d intervals, followed by a final 121-d finishing period. In the growing period, SHDL was added to the TMR which was mixed daily in a Calan data ranger (American Calan, Northwood, NH, USA). During finishing, SHDL was first pelleted with ground barley grain and then mixed into the TMR. The pellets were ground through a 2.0 mm screen and added to the finishing TMR daily. This approach prevented SHDL from separating from other dietary ingredients in finishing diet.
Orts were collected, weighed and dried weekly to determine daily feed intake. All cattle had free access to water throughout the experiment. The steers were weighed individually (unshrunk) using a single confinement livestock scale (Stathmas type 513417) on two consecutive days at the beginning and at the end of each feeding period and at 28-d intervals. Health status of the cattle was closely monitored by barn staff during the entire experiment.
Fat thickness was determined using ultrasound [22] at the end of the growing and the finishing periods. All steers were slaughtered commercially at the end of the finishing period. Carcasses were graded according to Canadian Beef Carcass Grading Regulations (www.cbef.com/beefquality.htm) and warm carcass weight, longissimus muscle area (LMA), marbling score and saleable meat yield were measured.
Laboratory analysis
Substrates, diets and orts were freeze-dried and ground to pass a 1.0-mm screen using a Wiley Mill (Model 4, Arthur H. Thomas Co., Philadelphia, PA, USA). Dry matter content was determined by oven drying (105°C for 48 h), and organic matter (OM) by ashing (550°C for 16 h). Total N was analyzed by flash combustion gas chromatography and thermal conductivity detection (NA1500, Carlo Erba Instruments, Rodano, MI, Italy). Neutral detergent fibre was determined as described by Van Soest et al [23] with α-amylase and sodium sulfite included in the analysis and was expressed as NDF inclusive of residual ash. Starch was analyzed as described by Herrera-Saldana et al [24]. Culture supernatants were assayed for VFA by gas chromatography and ammonia using a colorimetric method as described by Wang et al [16] and CH4 concentrations in headspace gas were determined by gas chromatography [25].
Calculation and statistical analyses
For in vitro experiments, GP and DMD were corrected using the blank control, and accumulations of VFA and ammonia were corrected for their concentrations at 0 h of incubation. CH4 production was calculated on per unit of DMD basis. Values from three (forage) or two (barley grain) vials from each treatment at each time point in each run were averaged and run was considered the experimental unit. For the feedlot experiment, DM intake (DMI), average daily gain (ADG), feed efficiency (expressed as ADG/DMI) and carcass characteristics were calculated using individual animal as the experimental unit.
All data were statistically analyzed by analysis of variance using the MIXED procedure of SAS [26]. Data for the in vitro experiments were analyzed for each diet as a complete randomized design with concentration of SHDL as a fix effect and run as random effect. Data in the barley grain experiment were analyzed as 3×3 factorial design with type of grain, SHDL concentrations and their interaction as main effects and run as a random effect. Linear and quadratic orthogonal polynomial contrasts were conducted to determine the effect of SHDL concentrations on fermentation measurements. The feedlot study was analyzed as a complete randomized design with treatment as a fixed effect and individual animal as a random effect. Significant differences among treatments in all experiments were tested using LSMEANS [26]. Significance was declared when p≤0.05, and trend was considered when p was between 0.05 and 0.10 in the animal experiment only.
RESULTS
Mixed forage
With mixed forage, SHDL linearly reduced (p<0.01) 48-h in vitro DMD, NDFD, CH4 production (mL/100 mg DM incubated) and acetate:propionate ratio (A:P; Table 1). However, CH4 production per unit of digested DM basis linearly increased (p<0.001) with the increase of SHDL concentrations. In contrast, GP, VFA production and the molar percentage of branch chained VFA (BVFA) were not affected by SHDL.

Effects of sodium hydroxide dehydrated lignin (SHDL) at 0, 0.25, 0.5, 1.0, and 2.0 mg/mL on ruminal fermentation of alfalfa/grass mixture after 48-h in vitro incubation
Barley grain
With barley grain, addition of SHDL linearly increased (p<0.001) 24-h DMD, but linearly reduced (p<0.001) CH4 production, NH3 accumulation (p<0.001), A:P (p<0.05) and molar proportion of BVFA (p<0.05; Table 2). However, CH4 production on per unit of digested DM basis was linearly decreased (p< 0.001) by SHDL. Addition of SHDL to barley grain did not affect GP or VFA production.
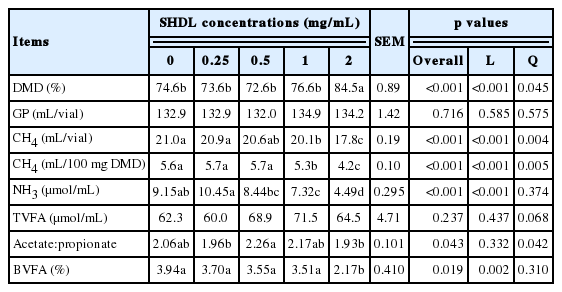
Effects of sodium hydroxide dehydrated lignin (SHDL) at 0, 0.25, 0.5, 1.0, and 2.0 mg/mL on ruminal fermentation of barley grain after 24-h in vitro incubation
There was no grain type by SHDL dosage interaction, so results were reported as main effects of grain type and SHDL (Table 3). As expected, 12-h DMD, GP and disappearance of starch all increased (p<0.001) with increasing severity of processing. However, additions of SHDL up to 2.0 mg/mL had no effect on DMD, but increased (p<0.05) GP at 12 h of incubation. Starch disappearance was also not affected by increasing concentrations of SHDL.

Effects of sodium hydroxide dehydrated lignin (SHDL) at 0, 0.5, 1.0, and 2.0 mg/mL on the 12 h in vitro ruminal fermentation of barley grain processed by different methods
Feedlot experiment
All cattle were of similar body weight at the beginning and at the end of the experiment regardless of level of SHDL supplemented (Table 5). Adding SHDL at 4 to 16 g/kg DM did not affect growth over the entire experiment. However, ADG tended to be linearly reduced (p = 0.090) in steers by SHDL during the growing, but not during the finishing period. Although there was no linear or quadratic response of feed intake to SHDL, steers fed the diet containing 16 g/kg of SHDL tended (p = 0.056) to have lower DMI than control steers in the finishing period. Feed intake over the entire experimental period tended to be linearly reduced (p = 0.100) with increasing level of SHDL in the diet, but there were no difference among treatments. Feed efficiency was similar among groups of steers during growing period regardless of the level of SHDL in the diet. However, during the finishing period the feed efficiency of steers responded quadratically (p = 0.059) to increasing levels of SHDL and at 8 g/kg DM of SHDL feed efficiency was improved as compared to Control (p = 0.038) steers. Over the entire experiment, no difference was observed in feed efficiency of steers among treatments. All steers were observed healthy and no veterinary attention or animal withdraw occurred during the entire experimental period.

Effects of sodium hydroxide dehydrated lignin (SHDL) on the growth performance of feedlot cattle fed barley grain and barley silage based total mixed rations (n = 15)
Most of the measured carcass traits were not affected by SHDL (Table 6). However, supplementation with SHDL at the dosages used in this study tended (p = 0.098) to linearly increase saleable meat. Carcass from steers fed diets with 16 g/kg DM of SHDL tended (p = 0.059) to have greater saleable meat, with fat cover tended (p = 0.076) to be less than that of control steers. Ultrasound measurements of LMA and average fat cover at the end of the experiment were consistent with carcass traits that were observed at slaughter. The LMA area measured at the end of growing period was similar among treatments, but tended (p = 0.09) to be linearly increased by SHDL at the end of the finishing period. At the end of growing period, the average fat cover was linearly (p<0.05) decreased by SHDL and steers fed SHDL at the level of 16 g/kg DM had lower (p<0.05) average fat cover than control steers.
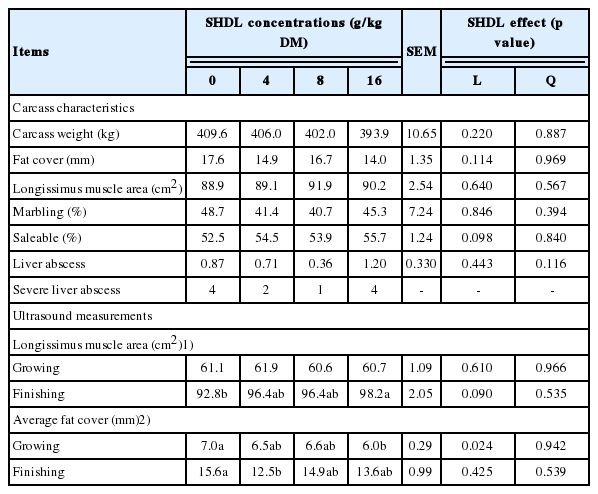
Effects of sodium hydroxide dehydrated lignin (SHDL) on the carcass traits of feedlot steers fed barley grain - barley silage growing and finishing diets (n = 15)
DISCUSSION
Effects of sodium hydroxide dehydrated lignin on ruminal fermentation
The linearly decrease of in vitro DMD for forage but a quadratic response of barley grain to SHDL suggests that the effects of SHDL on ruminal digestion may be diet dependent. With the forage diet, the SHDL mediated decrease of in vitro DMD corresponded with the decrease in NDFD. This suggests that SHDL inhibited fibrolytic activity, resulting in reduced fibre digestion and subsequent DM digestion in in vitro batch cultures. In contrast, SHDL had no effect on in vitro DMD and starch after 12-h, but increased 24-h DMD when barley grain was used as substrate. This suggests that unlike fibrolytic activity, SHDL did not inhibit the amylolytic activity of rumen bacteria in vitro.
These results are consistent with the observations that plant phenolics inhibit rumen cellulolytic bacteria and reduce fibre digestion [27,28]. Chemically, natural lignin is a complex polymer of phenyl propanoid subunits which occur in close physicochemical association with hemicellulose and cellulose. Hydroxyl, methoxyl, carbonyl, and carboxyl groups are the major chemical functional groups in lignin. During isolation, the physical and chemical links that exist between native lignin and other plant biopolymers are broken. However, there are chemical differences between native and commercially isolated lignin, such as reduction in molecular weight of the lignin, changes in ratio of functional groups and configuration of the molecule. Furthermore, some lignin extraction processes (but not the process that generates SHDL) also introduce sulfur-containing functional groups into the lignin molecule [15]. These structural changes likely introduce antimicrobial property that typically does not occur in native lignin. Antimicrobial activity, (especially anti-fibrolytic activity) of various phenolic compounds has been well recognized [28]. It is possible that the SHDL used in this study has similar antimicrobial properties that inhibit fibrolytic bacteria.
In vitro methane production per unit of mixed forage and barley grain substrates incubated linearly decreased with SHDL. This decrease in methane production suggests that SHDL may possess anti-methanogenic activity as has been reported for other naturally occurring plant compounds [29,30]. However, on a per digested DM basis, methane production from ruminal fermentation of the mixed forage diet linearly increased by SHDL, while it was linearly reduced with the grain diet. This indicates that SHDL may have more specifically inhibited methanogens with the grain diet, as starch digestion was unaltered, whereas with forage the decline in methane coincided with a reduction in NDF digestibility. Therefore, it is likely that the anti-cellulolytic activity of SHDL mainly contributed to the decreased methane production from the forage diet. Research has shown that plant bioactives reduce the major cellulolytic rumen bacterial populations and thereby decrease the amount of reducing equivalents available for methane production [29]. Compared to the forage diet, ruminal fermentation of barley grain produced much less methane on digested DM basis (9.5 vs 5.6 mL/g digested DM), which is consistent with the general trend that digestion of cellulosic feedstuffs produce more methane than feeds that consist primarily of starch [31]. In addition, the present study showed that SHDL decreased A:P as a result of increased propionate production from both forage and barley grain diets. This may have further decreased methane production as propionate is an alternative H sink to methane. Methane production has a negative correlation with energy utilization in ruminants [32]. Therefore, the decreased methane production by supplementing SHDL to grain diet suggests that SHDL may have the potential to improve energy utilization when animals are fed high grain diets. In addition, decreased A:P by SHDL would also contribute to improve energy efficiency. All of these indicate that inclusion of SHDL in finishing diet (high concentrate diet) may have the potential to improve feed efficiency.
Effects of sodium hydroxide dehydrated lignin on steer growth performance
Inclusion of SHDL at levels up to 16 g/kg DM in a barley based finishing diet resulted in a quadratic response in feed efficiency. However, this observation was not obtained for the growing diet, which contained a higher proportion of forage. In fact, in the growing period SHDL tended to reduce ADG. Therefore, the improvement in feed efficiency with SHDL observed at 8 g/kg DM in the finishing period reflects a reduction in feed intake without a corresponding reduction in ADG. These results were consistent with our previous work with the same SHDL in lambs [14]. It was shown that a reduction of DMI in lambs was mainly observed at 30 g of SHDL/kg DM and the lambs adapted to the dietary presence of SHDL and the feed intake returned to normal level after prolonged exposure. In the present study, the concentration of SHDL was much lower than that used in lambs yet the DMI of steers declined at 8 g SHDL/kg DM. This may be due to the difference in sensory sensitivity between cattle and sheep because it has been shown that cattle are able to detect tastes with greater sensitivity than sheep [33].
There is very little information on use of SHDL as a feed additive for ruminants. Phillip et al [13] reported a positive growth response in calves to another purified lignin product (Alcell extracted from hardwoods with ethanol) that was added to a grain-based diet at 12.5, but not at 25 or 50 g/kg DM. In their study, feed intake was not affected by the lignin, but feed efficiency was improved with 12.5 g lignin/kg diet DM. In contrast, inclusion of the same level of Alcell lignin in the diet of piglets or broiler chicks had no effect on growth, feed intake or feed efficiency [34,35]. However, it has been reported that supplementation of other lignin products improved growth performance of broiler chicks and geese [36,37]. The SHDL used in our study was derived from wheat straw after extraction with sodium hydroxide, suggesting that lignin from different sources may have different effects on animal performance. Our preliminary screening showed that antimicrobial activity differed among lignin products obtained from different plant sources and by different process (Wang et al., unpu-blished data).
Ultrasound measurements showed that SHDL linearly decreased fat cover during the growing period, but tended to linearly increase the LMA during the finishing period. These results are consistent with carcass measurements taken post-slaughter. The decreased fat cover during the growing period might be due to the decline in fibre digestion that usually produces more acetic acid for fat synthesis, a possibility that is supported by our observed decline in acetate production in vitro. Altogether, these results suggest that SHDL improved carcass quality, which is supported by the trend of increasing in the portion of saleable meat by SHDL although it is difficult to explain on a biological basis as to why such a big improvement in saleable meat yield. Nevertheless, the results of this study suggest that inclusion of SHDL at the level between 8 and 16 g/kg DM in diet containing high grain may benefit feedlot steers through potential in improving feed efficiency and carcass quality.
ACKNOWLEDGMENTS
This project was partially supported by Green Value SA and AAFC. Technical assistances from Long Jin, Zhong Xu, Allen Lu and feedlot barn staff are gratefully acknowledged. This is Lethbridge Research and Development Centre contribution number 38715072.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.