Association of Chicken Growth Hormones and Insulin-like Growth Factor Gene Polymorphisms with Growth Performance and Carcass Traits in Thai Broilers
Article information
Abstract
Molecular marker selection has been an acceptable tool in the acceleration of the genetic response of desired traits to improve production performance in chickens. The crossbreds from commercial parent stock (PS) broilers with four Thai synthetic breeds; Kaen Thong (KT), Khai Mook Esarn (KM), Soi Nin (SN), and Soi Pet (SP) were used to study the association among chicken growth hormones (cGH) and the insulin-like growth factor (IGF-I) genes for growth and carcass traits; for the purpose of developing a suitable terminal breeding program for Thai broilers. A total of 408 chickens of four Thai broiler lines were genotyped, using polymerase chain reaction-restriction fragment length polymorphism methods. The cGH gene was significantly associated with body weight at hatching; at 4, 6, 8, 10 weeks of age and with average daily gain (ADG); during 2 to 4, 4 to 6, 0 to 6, 0 to 8, and 0 to 10 weeks of age in PS×KM chickens. For PS×KT populations, cGH gene showed significant association with body weight at hatching, and ADG; during 8 to 10 weeks of age. The single nucleotide polymorphism variant confirmed that allele G has positive effects for body weight and ADG. Within carcass traits, cGH revealed a tentative association within the dressing percentage. For the IGF-I gene polymorphism, there were significant associations with body weight at hatching; at 2, 4, and 6 weeks of age and ADG; during 0 to 2, 4 to 6, and 0 to 6 weeks of age; in all of four Thai broiler populations. There were tentative associations of the IGF-I gene within the percentages of breast muscles and wings. Thus, cGH gene may be used as a candidate gene, to improve growth traits of Thai broilers.
INTRODUCTION
Poultry production is an important and diverse component of agriculture all over the world. Today, more attention has been given to indigenous animals in general, and poultry in particular; due to their quality of meat and sustainable production (Kaya and Yıldız, 2008). Meat from Thai native chickens is preferred by more Thai consumers than commercial broilers (Theerachai et al., 2003), due to their superior taste, meat texture, low fat and cholesterol, and high protein content (Promwatee and Duangjinda, 2010). However, the native chickens are inferior in production due to their low growth rates, as compared with commercial breed production. Promwatee et al. (2013) demonstrated that body weights of Thai synthetic chickens (50% native genetics) at 14 weeks of age, were between 1,532 to 1,561 g; which is significantly higher than the average body weight (1,280 g) of the typical 16 week Thai native chicken (Jaturasitha et al., 2008). Additionally, the market price of Thai native chickens is nearly two to three times higher than the commercial broiler (Wattanachant et al., 2004). Nowadays, hybrid chickens (with less than 50% native genetics) are more desirable for open-housing commercial production, due to the lower cost of production (faster growth) and greater tolerance to heat stress. Cross breeding of parent stock (PS) broiler sires with Thai synthetic breeds, in order to achieve a terminal hybrid of 75% broiler and 25% Thai native chicken (referred to as the Thai broiler), is of interest to the modern trait market. The products have a lower price, better taste, and better meat texture; compared to commercial broilers. In this regard, genetic improvements of parental lines for Thai broilers must be studied to meet the demands of consumers.
Growth performance and carcass traits are very significant economic traits in broiler production, and are controlled by sets of complex genes. Growth is a complicated procedure, regulated by a wide variety of neuroendocrine pathways (Zhang et al., 2008). For this reason, it is very difficult to make rapid progress using conventional methods of genetic selection within breeds (Zhang et al., 2008). Recent advances in molecular technology have provided new opportunities to evaluate genetic variability at the DNA level (Kaya and Yildiz, 2008). Therefore, the candidate gene approach has become a powerful technique for genetic improvement in the chicken breeding program. Applying a candidate gene may result in higher efficiency in detecting the desired traits necessary to improve production performance. The chicken growth hormone (cGH) and insulin-like growth factor-I (IGF-I) genes are among the most promising candidate genes for growth performance and carcass quality traits in chickens.
The cGH is a 22-kDa protein, containing 191 amino acid residues (Hrabia et al., 2008). In poultry, cGH consists of 4,101 base pairs, having five exons and four introns (Kansaku et al., 2008). Known as a polypeptide, hormone produced, and secreted by pituitary gland; cGH affects a variety of physiological functions in growth performance (Byatt et al., 1993; Apa et al., 1994). In the works of various authors, it was found that cGH gene is one of the most important genes affecting chicken performance traits, and plays a critical role in both growth and metabolism rates (Feng et al., 1997; Vasilatos-Younken et al., 2000).
IGF-I is known as one of the more predominant hormones necessary to support normal growth in chickens (Scanes, 2009; Boschiero et al., 2013). Furthermore, IGF-I is also involved in growth hormone secretion and regulation (Piper and Porter, 1997; Spencer et al., 1997; Rousseau and Dufour, 2007). In previous studies, the chicken IGF-I has been revealed to involve as many as 70 amino acids (Ballard et al., 1990). IGF-I is a complex system of peptide hormones that bind to the insulin-like growth factor I receptor (IGFIR), in order to activate their intrinsic tyrosine kinase domain activities (Denley et al., 2005). Additionally, the effect of IGF-I was observed on the protein synthesis of chicken embryo myoblast, cultured in a serum free medium (Kita and Okumura, 2000). Zhou et al. (2005) and Amills et al. (2003) reported that polymorphism of the IGF-I gene in the promoter and 5′-untranslated region (5′-UTR) was directly associated with chicken growth rate. There were dramatically higher IGF-I concentrations in the high growth rate line chickens, than those in the low growth rate line chickens (Beccavin et al., 2001).
To develop a suitable terminal breeding program it is necessary to study the relationship of cGH and IGF-I genes for use as candidate genes in Thai broilers. The purpose of the present study was to examine the association of cGH and IGF-I genes within the growth performance, and carcass traits in Thai broilers.
MATERIALS AND METHODS
Chicken populations
Four Thai broiler hybrids were established by crossing sires from a broiler breeder line (PS) with dams from four Thai synthetic chicken lines; namely, the Kaen Thong (KT), Khai Mook Esarn (KM), Soi Nin (SN), and Soi Pet (SP; Promwatee et al., 2013). A total of 408 individuals from the four Thai broiler lines were studied: PS×KT (n = 101), PS×KM (n = 104), PS×SN (n = 104), and PS×SP (n = 99). Phenotypic characteristics of all chicken lines are shown in Figure 1. All of four different colors of Thai synthetic dam lines are shown while only the white color of Thai broiler lines are shown due to the dominance of white color from PS broiler sire. The sample of Thai broiler chickens were supplied by the Research and Development Network Center for Animal Breeding, Khon Kaen University, Khon Kaen, Thailand. All chickens were fed ad libitum within the commercial broiler diet.

Phenotype characteristics of chickens in the mating program to produce Thai broiler. (A) Kaen Thong. (B) Khai Mook Esarn. (C) Soi Nin. (D) Soi Pet dam line. (E) Thai broiler male. (F) Thai broiler female.
Measurement of growth and carcass traits
Body weight (BW) of 408 chickens was recorded individually at hatching; and at 2, 4, 6, 8, and 10 weeks of age (BW 0, BW 2, BW 4, BW 6, BW 8, and BW10). The average daily gain (ADG) was calculated at two week intervals: 0 to 2 weeks of age (ADG 0–2), 2 to 4 weeks of age (ADG 2–4), 4 to6 weeks of age (ADG 4–6), 6 to 8 weeks of age (ADG 6–8), 8 to 10 weeks of age (ADG 8–10); as well as 0 to 6 weeks of age (ADG 0–6), 0 to 8 weeks of age (ADG 0–8), and 0 to 10 weeks of age (ADG 0–10). Description of data is described in Table 1 and 2. The formula of ADG was calculated using the equation below:

Descriptive statistics of data used in gene association study in PS×KT and PS×KM populations

Descriptive statistics of data used in gene association study in PS×SN and PS×SP populations
A total of 32 chickens were slaughtered at 10 weeks of age (8 chickens per line with 4 chickens per sex). All chickens were chosen as a representative sample based on average body weight and sex for each line. Carcass traits included live weight, dressing percentage, and the percentages of the measured breasts, drumsticks, wings, and thighs.
Genotyping with polymerase chain reaction-restriction fragment length polymorphism
Genomic DNA was extracted from the blood of 408 chickens. One mL of each individual blood sample was stored in a micro tube containing 100 μL of 0.5M ethylenediaminetetraacetic acid, as an anti-coagulant. Genomic DNA was isolated by using Guanidine Hydrochloride/Silica gel protocol (Goodwin et al., 2007).
The polymerase chain reaction (PCR) was performed in a 10 μL mixture containing 1 μL genomic DNA (50 ng), 1 μL 10× PCR buffer, 1 μL 2.5 μM of primers for each candidate gene, 1 μL 1 mM of dNTP (Thermo scientific, Waltham, MA, USA), 0.8 μL 25 mM MgCl2, and 0.1 μL 5U Taq DNA polymerase (RBC Bioscience, New Taipei, Taiwan). The primer characteristics of IGF-I (Zhou et al., 2005) and cGH (Nie et al., 2005) are shown in Table 3. PCR amplification was conducted under the following conditions: 95ºC for five minutes, followed by 30 to 35 cycles at 95ºC for 45 s, 58ºC to 68ºC for 30 to 45 s, and 72ºC for 30 to 45 s; followed by a final extension at 72ºC for five minutes.

Details of single nucleotide polymorphism markers and primers
Polymorphisms were detected by using the polymerase chain reaction-restriction fragment length polymorphism technique. The PCR products were digested in a total volume of 20μL of solution; containing 3μL of PCR product, 1 to 2 U of restriction enzymes, buffer, and H2O. The sample was then incubated at 37ºC overnight. Restriction patterns were visualized by 2% agarose gel electrophoresis, and stained in GelStar (GelStarInc, New York, NY, USA). Agarose gels were visualized and photographed under Gel Documentation System standards (SYNGENE, Madison, WI, USA).
Statistical analysis
Genotypic and allelic frequencies were calculated at each locus, as described by previous authors (Falconer and Mackay, 2001). Genotypes having a frequency lower than 2% were discarded from the analysis. The association of candidate genes and traits were analyzed with pooled data of four hybrids and adjusted line effect as fixed effect using the model below:
Where yijk is trait observation (BW and ADG), μ is overall population mean, Gi is the fixed effect of the genotype, Sj is the fixed effect of the sex, Hk is the fixed effect of the hatching, Cl is different hybrid cross effect, Cl×Gi is interaction effect between studied breed and gene, and eijk is the residual random error.
The association of candidate genes and traits were also analyzed separately for each hybrid cross using the following model:
Where yijk, μ, Gi, Sj, Hk, and eijk were described above. For carcass traits, according to the small number of samples, the association between candidate genes and traits were analyzed with pooled data from all hybrid cross, using the model as follow:
Where yijkl is a trait observation (carcass percentage), μ, Gi, Sj, Hk, Cl were described in previous model.
RESULTS AND DISCUSSION
Genotype and allele frequencies of cGH and IGF-I genes
Genotype and allele frequencies of cGH and IGF-I genes were calculated after genotyping the populations of four Thai broiler lines, as listed in Table 4. For the cGH gene, allele G is predominantly higher than allele A, in all four chicken populations. However, the AA genotype was counted with a frequency of 0.05 in the observations of PS×KM, and PS×SN populations. The AA genotype showed the lowest frequency (0.01) in the PS×SP population compared with the three other lines. For the IGF-I gene, the CC genotype was observed in all populations with lower frequencies (0.13 to 0.15) compared to that of the other genotypes.

Genotype and allele frequencies of cGH and IGF-I genes in Thai broilers
Association of cGH and IGF-I gene polymorphisms with growth traits
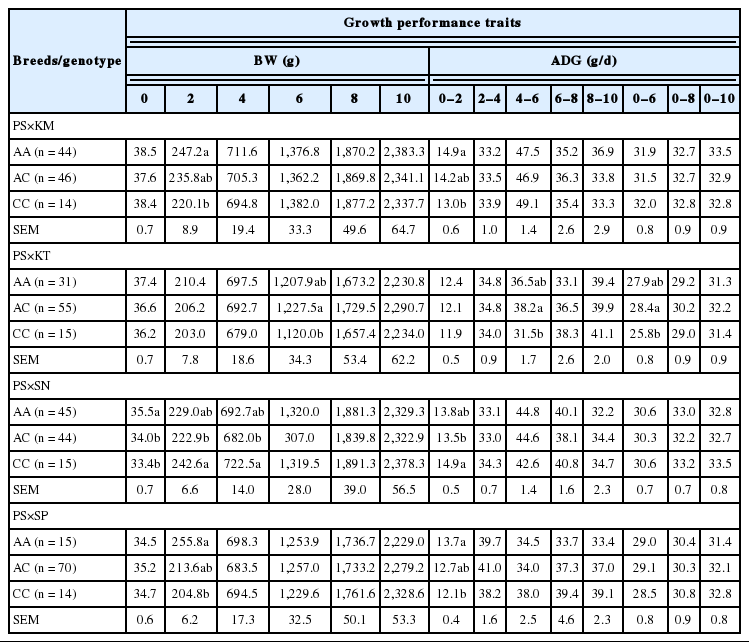
The analysis of association between the cGH and IGF-I gene polymorphisms and growth performance traits across all of four Thai chicken lines is shown in Table 5. A significant interaction between breed and gene was not found in almost all observed traits (p<0.05); except body weight at 4 and ADG at 2 to 4 weeks of age (p<0.05). The association of cGH gene was found in body weight trait at 4 and 6 weeks of age; and ADG at 2 to 4 and 0 to 6 weeks of age. Chicken with AG and GG genotypes showed higher BW and ADG (p<0.05) compared to that of the AA genotype. The analysis of the potential association between the cGH gene polymorphism and growth performance traits is summarized in Table 6. In the PS×KM hybrid, chickens with AG and GG genotypes demonstrated higher BW and ADG (p<0.01) compared to that of the AA genotype. Significant associations of cGH were found with only BW at hatching, and ADG 8–10 in PS×KT population (p<0.05). The cGH genotype effects (p>0.05) were not found in other hybrids (PS×SP and PS×SN).

Least square means of cGH and IGF-I genes on growth traits in Thai broilers across all chicken populations

Least square means of cGH gene on growth performance in Thai broiler populations
Nie et al. (2005) reported that the single nucleotide polymorphism (SNP) within the same region (G1705A) maintained a significant association with almost all growth traits, in an F2 reciprocal cross between the WRR and X Chinese chicken breeds. The allele A in the study by Nie et al. (2005) showed a positive effect on growth traits. However, our study concluded that allele G exhibited a generally positive effect on chicken growth and was completely dominant in all breeds (AG and GG having similar effects). These results confirm those found in the previous study of Thai native chickens (Chee), which revealed that all growth traits, including body weight and ADG, from 0 to 16 weeks of age; were significantly higher in the GG genotypes (Promwatee and Duangjinda, 2010).
The results of this study indicate that the cGH gene is associated with body weight and ADG in almost all recorded periods, within the PS×KM population. In this regard, the G1705A in intron 3 of cGH could have a direct effect on chicken growth performance by mediating cGH expression. Previous studies on other polymorphism in introns of the cGH gene also pointed out the association between chicken growth and carcass traits (Yan et al., 2003; Mehdi and Reza, 2012; Mu’in and Lumatauw, 2013). Hence, the cGH gene could be a potential marker for use in a marker-assisted selection programs. Further study of the associations between cGH and growth traits will be required to obtain more accurate results.
For IGF-I gene when the data was analyzed across all chicken populations (Table 5), the interaction between breed and gene was significant only at ADG during 0 to 2 weeks of age (p<0.05). There were significant associations of IGF-I gene with BW at 2 weeks of age; and with ADG at 0 to 2 and 4 to 6 weeks of age. Chickens with AA genotype showed higher body weight and ADG; except ADG during 4 to 6 weeks, while the AC genotype showed a higher result. The effects of polymorphism of the IGF-I gene on growth traits in Thai broilers is presented in Table 7. It was found that the IGF-I gene polymorphisms showed significant associations with only early periods of chicken growth in each of the four hybrid chickens (p<0.05). The IGF-I gene was chosen as a candidate gene to examine the associations of gene polymorphism in growth traits, within commercial broilers (Zhou et al., 2005; Kaya and Yıldız, 2008), synthetic breeds (Promwatee et al., 2013), and Thai native chickens (Chee) (Promwatee and Duangjinda, 2010). However, the association with the same region of IGF1-SNP1 in two chicken strains of the Black Penedesenca breed, showed only the association with ADG at 107d within a single strain (Amills et al., 2003). The results of this study indicate that the IGF-I gene had an effect on body weight and ADG in the early ages of all four hybrid crosses. However, the genotype effects were not clear. Therefore, this gene was regarded as still unsuitable for use as a marker for parental selection.
Least square means of IGF-I gene on growth performance in Thai broiler populations
The SNP variation in terms of allele substation effect against total genetic effect of cGH and IGF-I gene on growth performance traits across all chicken populations also was calculated and showed in Table 8. It was found allele A of cGH has negative effects for BW at most of ages from week 2 to 10. Chicken with AA had the lowest BW compared to other genotypes. The allele substitution effects are not clear for IGF-I.
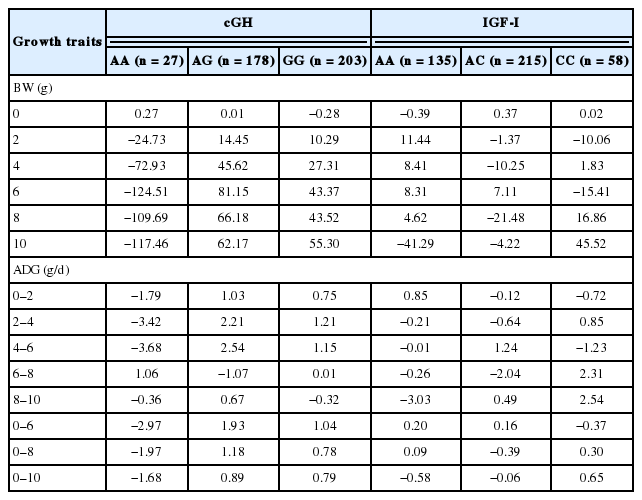
The single nucleotide polymorphism variation in terms of allele substitution effect against total genetic effect of cGH and IGF-I genes on growth traits across all chicken populations
Association of cGH and IGF-I gene polymorphisms with carcass traits
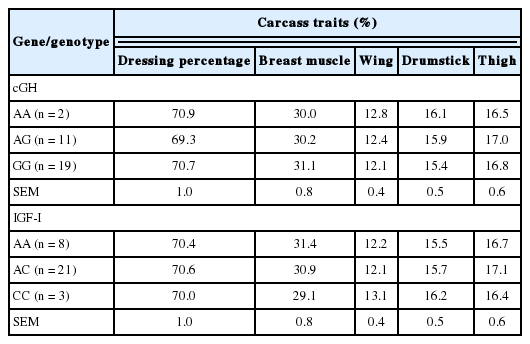
The probability values of the main effects of the cGH and IGF-I gene polymorphisms on chicken carcass traits are shown in Table 9. There was only tentative association within the four Thai broilers between the cGH and dressing percentage. A previous study revealed that the AA homozygote significantly differed from the GG homozygote in varied carcass traits in a F2 reciprocal cross between the WRR and X Chinese chicken breeds (Nie et al., 2005).
Least square means of cGH and IGF-I genes on carcass traits in Thai broilers
Similarly, in IGF-I gene polymorphism, there was no significant association with any carcass traits. Nevertheless, tentative associations with the breast and wing percentages were found (p<0.15). Previous study of the same mutation of IGF-I in Thai synthetic chicken lines reported that a significant association of IGF-I was found in only dressing and pectoralis major weight percentages in a single line (Promwatee et al., 2013). Otherwise, the same region of IGF-I within our study revealed that there were significant associations with all observed carcass traits, in F2 Leghorn and Fayoumi cross chickens; at eight weeks of age (Zhou et al., 2005). The opposite effect seen in our study may be the result of chicken samples having been selected at different ages and of different genetic backgrounds.
CONCLUSION
This study found some significant effects of cGH and IGF-I SNP associated with chicken growth traits. However, the effects, though significant, could not generally be used across breeds. The selected cGH genotypes AG or GG, suggest breeding KM female with male PS chickens, in order to create the PS×KM hybrid, produces a better growth performance and has a greater potential to develop into Thai broilers. The IGF-I gene polymorphisms did not suggest a similar potential, as the genotype effects were unclear. In summary, the cGH gene polymorphisms may be used as genetic markers for improving growth traits in breeding programs for commercial hybrid chickens.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the Research and Development Network Center for Animal Breeding (Native Chicken), Khon Kaen University for help in managing chickens and collecting data. The authors are most grateful to Department of Animal breeding and Genetics, Institute of Animal Science for Southern, Vietnam and the Vietnam Ministry of Education and Training, and Vietnam Ministry of Agriculture and Rural Development for granting the Ms. C. scholarship and for the research funding. The authors wish to acknowledge Animal Genomic Laboratory, Faculty of Agriculture for providing research facilities. This work was also supported by Higher Education Research Promotion and National Research University Project of Thailand, and the Office of the Higher Education Commission, through the Food and Functional Food Research Cluster of Khon Kaen University.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.