Effects of Dietary Fermented Chlorella vulgaris (CBT®) on Growth Performance, Relative Organ Weights, Cecal Microflora, Tibia Bone Characteristics, and Meat Qualities in Pekin Ducks
Article information
Abstract
Fermented Chlorella vulgaris was examined for its effects on growth performance, cecal microflora, tibia bone strength, and meat qualities in commercial Pekin ducks. A total of three hundred, day-old male Pekin ducks were divided into three groups with five replicates (n = 20 ducklings per replicate) and offered diets supplemented with commercial fermented C. vulgaris (CBT®) at the level of 0, 1,000 or 2,000 mg/kg, respectively for 6 wks. The final body weight was linearly (p = 0.001) increased as the addition of fermented C. vulgaris into diets increased. Similarly, dietary C. vulgaris linearly increased body weight gain (p = 0.001) and feed intake (p = 0.001) especially at the later days of the feeding trial. However, there was no C. vulgaris effect on feed efficiency. Relative weights of liver were significantly lowered by dietary fermented C. vulgaris (linear effect at p = 0.044). Dietary fermented C. vulgaris did not affect total microbes, lactic acid bacteria, and coliforms in cecal contents. Finally, meat quality parameters such as meat color (i.e., yellowness), shear force, pH, or water holding capacity were altered by adding fermented C. vulgaris into the diet. In our knowledge, this is the first report to show that dietary fermented C. vulgaris enhanced meat qualities of duck meats. In conclusion, our study indicates that dietary fermented C. vulgaris exerted benefits on productivity and can be employed as a novel, nutrition-based strategy to produce value-added duck meats.
INTRODUCTION
Chlorella vulgaris (C. Vulgaris) is a genus of single-cell green algae belonging to the Phylum Chlorophyta and has been widely acknowledged as a functional food worldwide. It is rich in protein with balanced essential amino acid profiles, lipid, carotenoids, vitamins and minerals (Kay, 1991). Due to its rapid multiplication, it is considered as a useful protein candidate among future food resources (Kang et al., 2004). Earlier, intact C. vulgaris was known to have low protein digestibility, which was mainly attributed to the presence of rigid cell walls (Shelef and Soeder, 1980). However, protein digestibility of C. vulgaris fermented with beneficial bacteria or yeasts was dramatically improved in rats (Buckenhüskes et al., 1990; Keijiro, 2011). In addition to the use of the protein resource for humans, C. vulgaris was also tested as a replacement of antibiotics in food-producing animals (Yan et al., 2012). Dietary C. vulgaris was reported to possess immunemodulatory activity in broiler chickens (Kotrbacek et al., 1994), to increase growth performance in pigs (Kim, 2011; Yan et al., 2012), and to enhance egg quality in laying hens (Zheng et al., 2012). Despite there being several experiments investigating fermented C. vulgaris in broiler chickens, laying hens and pigs, the utilization of fermented C. vulgaris as a duck feed additive has never been reported. In the present study, we investigated the effects of fermented C. vulgaris on growth performance, meat qualities, cecal microflora, relative weights of organs, and tibia bone characteristics in Pekin ducks.
MATERIALS AND METHODS
Preparation of fermented Chlorella vulgaris
In this study, a commercially available fermented C. vulgaris product (CBT®, Celltech, Co. Ltd, Eumsung, Korea) was used which was produced by the patent fermentation process as described earlier (Zheng et al., 2012). Briefly, C. vulgaris was inoculated with baker’s yeast and lactic acid bacterium. After 72 h of dry fermentation with cereal broth, fermented C. vulgaris was then dried at low temperature and ground into powder. The analyzed chemical contents of the CBT® were as following; crude protein (CP), 24.85%; crude fiber, 5.36%; ether extract, 2.42%; Ca, 0.20%; and P, 0.65%.
Experimental design, animals and management
Three hundred day-old male Pekin ducks were divided into three dietary groups with five replicates (n = 20 ducklings per replicate) in a completely randomized design model. The control basal diet was formulated to meet or exceed the nutrient recommendations of the NRC (1994) and Korean feeding standard for poultry (2012). The starter basal diet in a crumble form was fed from hatch to 3 wk, and contained 22% CP and 2,950 kcal nitrogen corrected true metabolizable energy (TMEn)/kg of diet. The grower basal diet in a pellet form was provided from 3 to 6 wk, and had 18% CP and 3,100 TMEn/kg of diet. The formula and chemical compositions of the experimental diets are shown in Table 1. The supplement CBT® was added into the basal diet to the inclusion rate of 0, 1,000, or 2,000 mg/kg. The diet and water were freely available. The ducklings were initially reared at 33°C, and the room temperature was gradually decreased by 4°C weekly to reach 22°C and kept constant thereafter. They were housed on floor pens with rice husks as a bedding material with a 23 h light and 1 h dark cycle throughout the experimental period. All animal care procedures were approved by the Institutional Animal Care and Use Committee at Konkuk University.

Ingredients and compositions of the experimental diets
Sampling and measurements
The body weight (BW) and feed intake on a pen basis were recorded weekly and used to calculate gain:feed ratio. At the end of experimental period, eight ducks per pen were randomly selected, weighed individually and euthanized by cervical dislocation. At necropsy, right breast and leg meats, liver, spleen and abdominal fat were immediately removed, blotted onto dry towel, weighed and expressed as relative weights per 100 g of BW.
For microbial test, cecal content per bird was aseptically sampled into preweighed sterile tubes and homogenized followed by serial 10-fold dilution with sterile phosphate buffered saline as described elsewhere (Miller and Wolin, 1974). Bacterial total microbes were enumerated on nutrient agar (Difco, BD science, Detroit, MI, USA), presumptive lactic acid bacteria on MRS agar (Difco, BD science, Detroit, MI, USA) and presumptive coliform bacteria on MacConkey agar (Difco, BD science, Detroit, MI, USA). All inoculated plates were incubated aerobically at 37°C for 24 h. The results obtained were presented as base-10 logarithm colony-forming units (cfu) per gram of cecal content (Tuohy et al., 2002).
To determine the cooking loss, 60 g of fresh breast meat was boiled in polyethylene bag, immersed in 80°C water bath for 30 min, and cooled to room temperature for 30 min. The cooking loss was calculated as the difference between the uncooked and cooked meat (Bouton et al., 1971). The water holding capacity was estimated according to the filter paper pressed method (Grau and Hamm, 1953). Approximately, 300 mg of breast meat was weighed on a Whatman filter paper (No. 2, UK) and the sample was pressed between two plexiglass plates for 3 min. The areas of pressed sample and water were measured using planimeter (Type KP-21, mitutoyo, Kawasaki-shi, Japan). The pH of breast meat was measured in triplicate with a pH meter (Model 340, Mettler-Toledo, Urdorf, Switzerland). Briefly, 1 g of breast meat was cut into small pieces, homogenized with 9 mL of distilled water for 1 min in an Ultra-Turrax (Model No. T25, Janke and Kunkel, Staufen, Germany). The instrumental colors of fresh meat, including lightness (L*), redness (a*) and yellowness (b*), were measured by a reflectance colorimeter (CR 210, Minolta, Tokyo, Japan) using illuminant source C. Color was measured in triplicate on the bone-side surface of each sample. The colorimeter was calibrated using a standard white ceramic tile.
The right tibia (meat removed) was used to analyze the breaking strength using an Instron Materials tester (model 3342, Instron Corp., Canton, MA, USA) with Automated Materials Test System software version 4.2. The weights, diameters, and lengths of right tibia were measured. Tibia diameters were calculated as the mean value of both narrow and wide parts. The deformation rate was 5 mm/min. Tracing of force was recorded at a constant rate. The graphs showed the plateau curve of applied maximal force (KN) to measure the bone strength as expressed as energy stored in the bone. After measuring the bone breaking strength, the bones were dried at 105°C, weighed, ashed in a muffle furnace at 600°C for 24 h, cooled in a desiccator, and weighed again.
Statistical analysis
The experimental data were analyzed using Generalized Linear Model procedures of SAS (SAS Institute, 2002). Orthogonal polynomials contrasts were used to determine the linear and quadratic effects of the dietary CBT® supplementation according to the following general model: Y = μ+α+ɛ, where Y was the observed response variables; μ was the overall mean; α was the effect of diet and ɛ was the random error. The pen was considered as an experimental unit. An alpha level of 0.05 was considered the statistical significance.
RESULTS AND DISCUSSION
Growth performance
The effect of dietary fermented C. vulgaris on growth performance is presented in Table 2. The final BW was linearly (p = 0.001) increased as the inclusion rate of fermented C. vulgaris into diets increased. Similarly, dietary C. vulgaris linearly increased the BW gain (p = 0.001) and the feed intake (p = 0.001), especially from 1 to 42 days post-hatch. However, there was no significant effect on feed efficiency between the treatment and control groups. Thus, it is concluded that the increased BW in C. vulgaris-treated ducks is the consequence of the increase in feed intake, but not by efficient nutrient utilization. It has been suggested (Zheng et al., 2012) that fermentation of C. vulgaris may improve its flavor as the underlying mechanism of increased feed intake in laying hens. However, it is well understood that taste sensors in poultry are less developed compared with those of the mammals. Thus, further study is needed to clarity the increased feed intake by fermented C. vulgaris in Pekin ducks as seen in this study.

Effect of fermented Chlorella vulgaris (CBT®) on growth performance in Pekin ducks
Relative weights of various organs
The effect of dietary fermented C. vulgaris on relative weights of the various organs and tissues are presented in Table 3. Relative weights of liver were significantly lowered by dietary fermented C. vulgaris (quadratic effect at p = 0.091). At this stage, a clear explanation of the decreased relative weights of liver in response to feeding fermented C. vulgaris is unknown and requires further study. Our finding is in contrast to the previous study (Zheng et al., 2012) which showed no effect of C. vulgaris at the inclusion rate of 1,000 or 2,000 mg/kg in diet on the relative weights of liver in 80-wk-old laying hens compared with those of the control group. The difference in the relative liver weight between our study and Zheng et al. (2012) could be the breed difference as we used the rapidly growing meat-type duck while the latter used slow-growing laying hens. On the other hand, spleen, abdominal fat, and the breast and leg muscles of ducks when adjusted to 100 g of BW were not affected by supplementation of the fermented C. vulgaris.
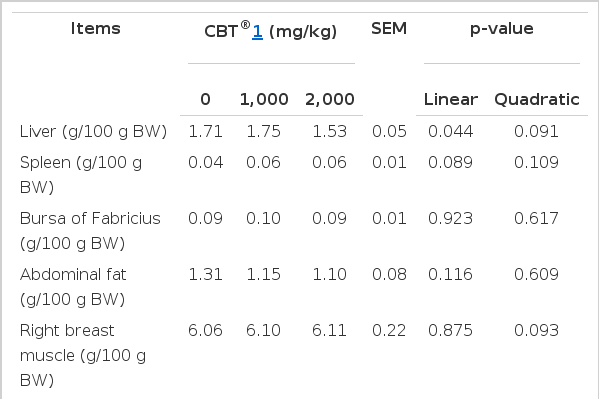
Effect of fermented Chlorella vulgaris (CBT®) on relative weights of various tissues in Pekin ducks
Cecal microflora
The findings by Pratt et al. (1944) who reported that chlorellin, the active component in Chlorella, has an antibiotic effect, and by Amaro et al. (2011) who reported that methanol extracts of C. vulgaris lowered E. coli and Salmonella, prompted us to measure the population of cecal microflora in this study. As shown in Table 4, dietary fermented C. vulgaris did not affect total microbes and lactic acid bacteria in cecal contents (Table 4). However, the population of cecal coliform bacteria in ducks fed diet with 2,000 mg/kg fermented C. vulgaris tended to be lower compared with their control-diet fed counterparts (linear effect at p = 0.064), indicating that C. vulgaris may have a positive effect on improving cecal microflora. Janczyk et al. (2009) reported that when 23 wk-old brown-egg laying hens were fed with powder and pellet forms of C. vulgaris at 5 g/kg of diet, the number of Lactobacilli spp. increased in the crop and the ceca. In this study, we did not attempt to measure the active components such as chlorellin, which is known to have the antibiotic effect (Pratt et al., 1944) or biomolecules which might be produced during the C. vulgaris fermentation process with baker’s yeast and lactic acid bacterium. It should be pointed out that the concentrations that we used in this study would be sufficient to exhibit the biological effect, if any, on cecal microbiota. In any events, a clear explanation regarding the lack of effect by dietary fermented C. vulgaris on cecal microbiota is not readily available.

Effect of fermented Chlorella vulgaris (CBT®) on cecal microflora in Pekin ducks
Physicochemical properties of tibia
Dietary fermented C. vulgaris did not affect tibia length (Table 5), but it significantly increased breaking strength of the tibia (linear effects at p = 0.001 and quadratic effects at p = 0.01). Although not significant (linear p = 0.080), dietary fermented C. vulgaris numerically increased ash content in tibia. Calcium metabolic analysis can be measured by either compressive strength, ash content, mineral content, or bone density measurement (Crenshaw et al., 1981; Garlich et al., 1982; Akpe et al., 1987; Watkins and Southern, 1992). Thus, increased tibia strength as seen in this study might be related to the enhanced calcium storing capacity of the tibia in ducks fed diets containing the fermented C. vulgaris. In humans, short- or long-term intake of Chlorella increased bone density and bone metabolism (Kim et al., 2002), which may support our latter assumption on C. vulgaris and calcium metabolism. In any event, further studies on apparent metabolizabilities of calcium and/or phosphorus could explain the observed effect of dietary fermented C. vulgaris on the calcium utilization in Pekin duck.
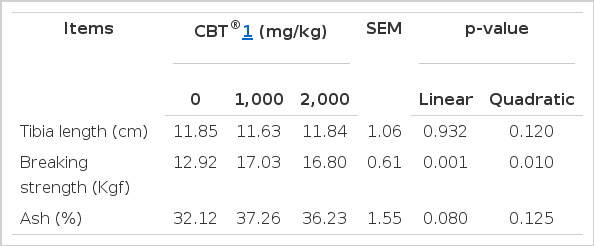
Effect of fermented Chlorella vulgaris (CBT®) on physicochemical properties of tibia in Pekin ducks
Meat quality
It is reported that C. vulgaris contains carotenoids (Kay, 1991), which are well-known as coloring agents in skin pigmentation. Thus, various meat quality parameters were measured including meat color intensity, shear force, pH, cooking loss and water holding capacity. The effect of dietary fermented C. vulgaris on meat quality is presented in Table 6. Dietary C. vulgaris linearly increased (p = 0.012) breast meat yellowness, but no effect on lightness or redness was monitored. In leg meats, dietary C. vulgaris significantly increased meat lightness (linear effect at p = 0.002) and yellowness (quadratic effect at p = 0.008). The shear force (p = 0.048) and pH (p = 0.004) of breast meats, but not leg meats were linearly increased by C. vulargis. The cooking loss of the breast or leg meats was not affected by dietary C. vulgaris. Finally, water hold capacity was increased quadratically (p = 0.044) in breast meat, but not in leg meat, when ducks were fed diets supplemented with C. vulgaris.
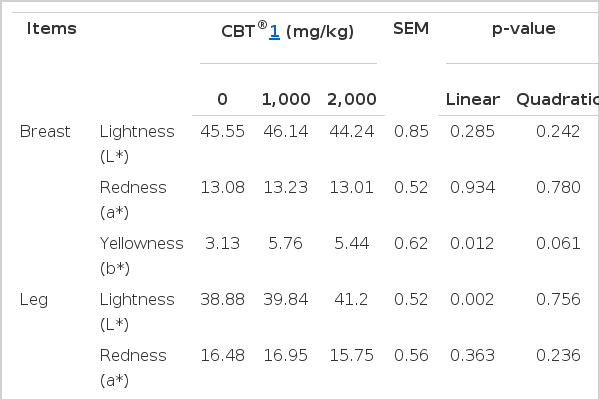
Effect of fermented Chlorella vulgaris (CBT®) on meat qualities in Pekin ducks
As expected, the yellowness of duck meats were substantially increased as C. vulgaris increased in diets, indicating efficient transfer of active carotenoids present in C. vulgaris. In general, poultry skin pigmentation (i.e., yellowness) can be increased by feeding natural or synthetic pigments such as carotenoids in diets (Castaneda et al., 2005), which was the case in this study. According to the study by Park et al. (2005), the quality of edible meat can be determined by pH, which is associated with water retention capacity and color. Indeed, we also observed that the pH and water hold capacity of the breast meat increased as C. vulgaris increased in diet. In our knowledge, this is the first report that fermented C. vulgaris enhanced duck meat qualities.
CONCLUSIONS
In conclusion, these results indicated that dietary fermented C. vulgaris increased growth performance, tibia bone strength, and enhanced meat qualities in Pekin ducks. It is demonstrated that fermented C. vulgaris has strong potentials to produce value-added duck meats and can be introduced as a novel feed additive in duck industry.
ACKNOWLEDGMENTS
This work was supported by the Konkuk University, 2013.