Nutritional and Hormonal Induction of Fatty Liver Syndrome and Effects of Dietary Lipotropic Factors in Egg-type Male Chicks
Article information
Abstract
This experiment was conducted with male chicks to investigate the influence of hormones and nutrients on the development of fatty liver syndrome (FLS) as well as the effects of dietary lipotropic factors on hepatic fat accumulation and lipogenic enzyme gene expression. A total of two-hundred sixteen 4-wk-old Hy-Line male chicks were divided into six groups and fed an experimental diet (T1, low-energy diet with low levels of lipotropic factors; T2, high-energy diet with low levels of lipotropic factors; T3 and T5, low-energy diet with high levels of lipotropic factors; T4 and T6, high-energy diet with high levels of lipotropic factors) for six weeks. The chicks in T5 and T6 groups were treated with intramuscular injections of estradiol benzoate for three days prior to biopsy and clinical analysis of FLS. Chicks treated with estrogen had significantly greater liver weights than untreated chicks. The abdominal fat contents were increased in chicks consuming high-energy diets as compared to those consuming low-energy diets. Treatment with estrogen significantly increased the concentrations of serum cholesterol, triacylglycerol and phospholipid (p<0.05). The hepatic triacylglycerol levels were tenfold higher in the estrogen treated chicks than in the untreated chicks. There were no significant differences in malondialdehyde levels between the treatment groups. Estrogen treatment dramatically increased the levels of fatty acid synthetase, acetyl-CoA carboxylase and ApoB mRNA. The results indicated that treatment with exogenous estrogen in growing male chicks induced hepatic fat accumulation, which might be partially due to increased lipogenic enzyme gene expression.
INTRODUCTION
FLS and fatty liver hemorrhagic syndrome (FLHS) are caused by diverse nutritional and metabolic factors (Butler, 1976; Jensen, 1976). Genetic, environmental, and hormonal factors are causative of these metabolic diseases in poultry (Squires and Leeson, 1988; Hansen and Walzem, 1993). An interaction between these factors may contribute to FLS and FLHS development; however, earlier studies have been unable to elucidate the exact cause of FLHS.
Previous work has investigated whether high energy diets result in FLHS following fat accumulation. Indeed, excessive energy intake regardless of ingredients such as glucose, starch, and corn oil caused FLHS in 28-wk-old laying hens force-fed over the 150% of normal feed intake over a prolonged period (Polin and Wolford, 1976). In contrast, an earlier study found that level of energy intake did not correlate with FLHS (Ivy and Nesheim, 1973). Excessive energy intake in these hens elevated hepatorrhagia (Polin and Wolford, 1977). On the other hand, in hens treated with exogenous estradiol benzoate, there was an elevation in plasma lipid concentration but not in hepatic lipid concentration (Akiba et al., 1983). Because estrogen treatment increased hepatic lipid excretion rather than accumulation.
Lipotropic factors, such as methionine, choline, and vitamin E are used to treat or prevent FLS and FLHS. Methionine and choline deficient diet results in FLS, because it causes increased oleic acid absorption and decreased triacylglycerol secretion in the liver (Rinella et al., 2008). Diets containing choline, vitamin B12, and vitamin E decreased hepatic triacylglycerol accumulation in laying hens (Couch, 1956).
Estrogen concentration of plasma was controlled by various diets (Akiba et al., 1982). Diets with extra-volume of choline, vitamin B12, and vitamin E were shown to reduce FLHS after force-feeding (Wolford and Polin, 1975). In other study, water containing high concentrations of Ca, Mg, Sr, Na, Fe, and Ba caused FLHS in laying hens (Jensen et al., 1976). There was no correlation between water including Mn and FLHS, and diet mainly composed of corn had no beneficial effect on FLHS. Hepatic fat concentration and hemorrhage were elevated in hens fed a diet containing low level of Ca (Roland et al., 1985). These hens also showed symptoms of ovarian hypertrophy, possibly due to injection with endogenous estrogen. Therefore, although numerous causative factors of FLS and FLHS have been identified, reports have not yielded consistent results.
Immature hens challenged with exogenous estrogen were used mainly as a FLS induction model; however, it produced change of hepatic lipid metabolism due to estrogen administration (Dashti et al., 1983). Using this method, estrogenized male chicks exhibited a 6-fold increase in hepatic fat concentration, likely due to increased triacylglycerol (Hermier et al., 1996). In vitro, livers from hens treated with estrogen displayed a marked increase in fatty acid synthesis and subsequent lipid accumulation (Rosebrough et al., 1989).
The objective of the present study was to determine the effects of dietary energy levels, lipotropic factors and estrogen treatment on i) growth performance ii) the concentrations of various lipid fractions in serum and liver and iii) mRNA expression of ApoB related to lipid metabolism in immature laying-type male chicks.
MATERIALS AND METHODS
Birds and experimental design
Two-hundred sixteen 4-wk-old Hy-Line male growing chicks were assigned to six treatments according to a completely randomized design. Each treatment had three replications with 12 birds per cage. The chicks were housed in wire cages (1.8×1.8×2.0 m), and each cage was equipped with a feeder and a water trough. The birds had ad libitum access to feed and water for six weeks. During the experimental period, the lighting was on throughout the day. The room temperature was 21±1°C.
The treatments included: i) T1: TMEn level of 11.72 MJ/kg, low level lipotropic factors, and no estradiol benzoate injection, ii) T2: TMEn level of 12.89 MJ/kg, low level lipotropic factors, and no estradiol benzoate injection, iii) T3: TMEn level of 11.72 MJ/kg, high level lipotropic factors, and no estradiol benzoate injection, iv) T4: TMEn level of 12.89 MJ/kg, high level lipotropic factors, and no estradiol benzoate injection, v) T5: TMEn level of 11.72 MJ/kg, high level lipotropic factors, and 2 mg/kg BW injection of estradiol benzoate, vi) T6: TMEn level of 12.89 MJ/kg, high level lipotropic factors, and 2 mg/kg BW injection of estradiol benzoate. TMEn level of 11.72 MJ/kg in the diet was applied to growing chicks of brown-egg-laying strain. The 11.72 MJ/kg applies to low energy, and 12.89 MJ/kg of high energy is 10 percent higher than low energy. The low level of lipotropic factors was no shortage of vitamin requirement presented in NRC. However, the high level of lipotropic factors contained 20,000 IU of vitamin A, 4,000 IU of vitamin D3, 10.5 IU of vitamin E, 4 mg of vitamin K3, 4 mg of vitamin B1, 12 mg of vitamin B2, 6 mg of vitamin B6, 0.02 mg of vitamin B12, 0.1 mg of biotin, 60 mg of niacin, 20 mg of pantothenic acid, and 2.0 mg of folic acid. Estradiol benzoate was injected into the pectoral muscle during the last three days of the experiment.
The experimental diet (Table 1) consisted primarily of corn, soybean meal, and wheat bran was supplemented with vitamins and minerals to meet National Research Council (NRC, 1994) requirement. Body weights and feed intake were recorded weekly. Wasted feed was collected and weighted to precisely evaluate feed intake. Then, weight gain and feed conversion ratio were calculated. Ten birds from each treatment were slaughtered at wk 6. All animal care procedures were approved by the Institute Animal Care and Use Committee at Konkuk University. The liver and abdominal fat weight were measured and converted into relative weights per 100 g of body weight.
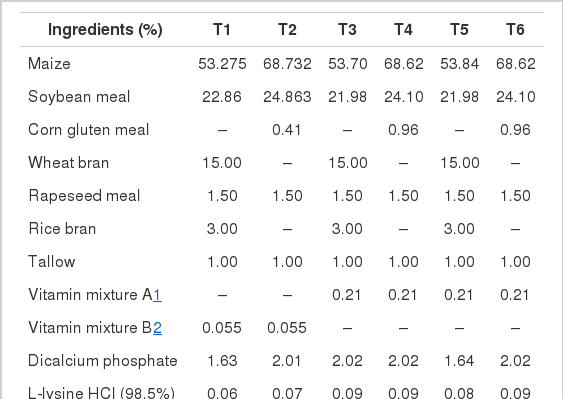
The ingredients and chemical compositions of experimental diets
Serum collection and chemical analysis
At the end of experimental period, 10 birds from each treatment were randomly selected and individually weighed. The blood samples were collected via wing vein using sterilized syringes for determination of the various blood profiles. At necropsy, the liver and abdominal fat were immediately removed and weighed. The activities of glutamic-oxaloacetic transaminase (GOT) and glutamic-pyruvate transaminase (GPT) were measured by a colorimetric method using the GOT-GPT assay kit (BCS GOT-GPT assay kit, Bio Clinical System Corporation), according to the manufacturer’s instructions.
Determination of the lipid fraction in liver and serum
The modified method was used to determine the total lipid fraction of the liver and serum (Folch et al., 1957). The contents of each lipid fraction were separated by thin layer chromatography (TLC) on silica gel chromarods (Chromarod-S III, Mitsubishi Kagaku Iatron, Inc., Japan), an exclusive media in the same manner of normal phase TLC. Rods were developed in hexane:diethyl ether:formic acid (60:10:0.1 ml). And then, each lipid fraction was quantified using the IATRO SCAN (TH-10 TLC/FID analyzer, Iatron, Ltd., Japan) technique as previously described (An et al., 1997).
Measurement of malondialdehyde concentration
Lipid oxidation was determined the thiobarbituric acid reactive substance (TBAR) values in the liver using a modified method (Botsoglou et al., 1994). To estimate the malondialdehyde (MDA) contents in the liver, a 1.5 g sample was weighed in a centrifuge tube and thoroughly homogenized with 8 ml of 5% aqueous trichloroacetic acid and 5 ml of 0.8% butylated hydroxytoluene in hexane. The mixture was centrifuged for 3 min at 3,000 g. After a chemical reaction with 8 ml of 2-thiobarbituric acid, the MDA concentration was analyzed by third derivative spectrophotometry against a blank reaction mixture (Beckman DU-530, Beckman Counter, Inc., Fullerton, USA). Measurements were conducted in duplicate. The liver color from 10 birds per treatment was scored by three investigators. The color scored from 1 (dark red) to 5 (light yellowish red). The standard for liver color score is shown in Figure 1.
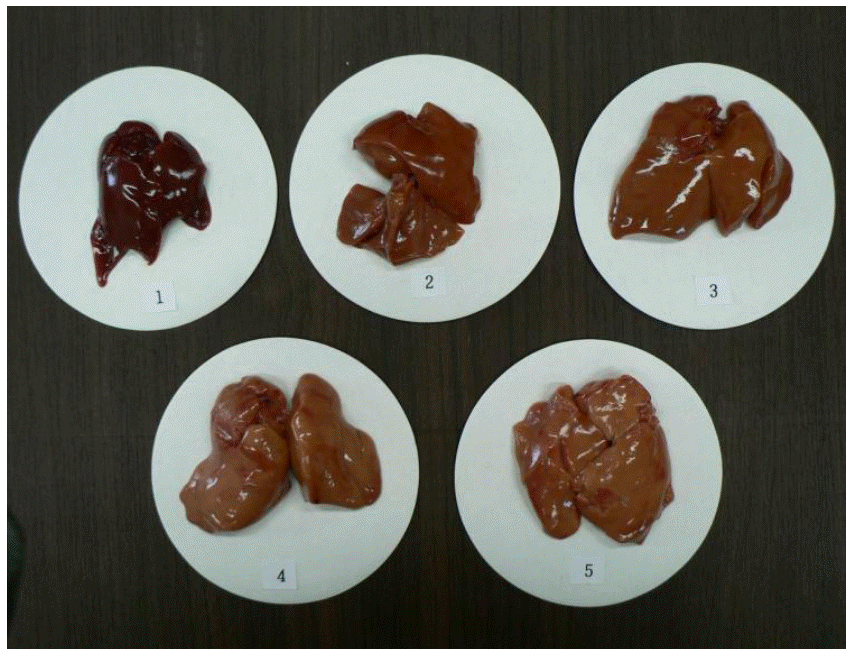
Standard for liver color score. The color of normal liver was scored 1 and the color score from 2 to 5 (from dark red to light yellowish red) was judged by three investigators.
Quantitative real-time RT-PCR analysis
Total RNA was extracted from liver samples using a modified acid/guanidinium/phenol/chloroform method (TRIzol reagent, Invitrogen, USA) under RNase free conditions (Chomczynski and Sacchi, 1987). DNase inactivation was conducted by chemical reaction in order to protect DNA, and complementary DNA (cDNA) was synthesized using Accupower® RT-PreMix (Bioneer, Daejeon, Korea). The primer sequences of the housekeeping gene are presented in Table 2. PCR was carried out with the synthesized cDNA as a template. The mixture was as follows: 0.5 ml of Taq polymerase (5 units/μl, Takara EX Taq™, Takara EX Taq™ polymerase, Takara, Japan), 2 ml of dNTP mixture (2.5 mM, Takara, Japan), 1 mg of template, 1 mg of each of primer (25 pmole), 13.5 mg of tertiary distilled water. After the DNA amplification, DNA marker and electrophoresis were conducted. Then, the optical density (OD) values of the β-actin DNA band and those of FAS, ACC, and ApoB were measured by an image analyzer (Biocapt ver. 99.4, Vilber Loumat, France).
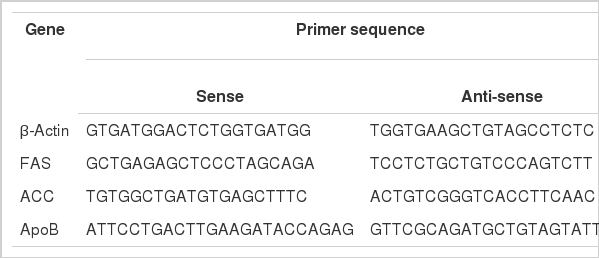
Primer sequences of β-actin, FAS, ACC, and ApoB
Statistical analysis
The experimental data were analyzed by comparing means according to Duncan’s multiple range test (Duncan, 1955) using the generalized linear model procedures of SAS (SAS, 2002). Statistical significance was defined as p<0.05.
RESULTS AND DISCUSSION
Growth performance and relative weight in liver and abdominal fat
The effects of dietary energy content, lipotropic factors, and estradiol benzoate injection on growth performance and relative weight in liver and abdominal fat are presented in Table 3. There were no statistically significant differences in final body weight and daily weight gain, but daily weight gain was lowest in T5. Daily feed intake did not show the any significant differences statistically. However, T5 showed significantly higher feed conversion rate (FCR) than the other groups consuming low energy diets (p<0.05). This may have been due to intramuscular stress from the injection. But FCR of T6 injected with equal volume of estrogen was similar compared to the other groups, which was considered to be effect of the high energy diet.

In previous study, feed intake increased in the growing chicks challenged with estrogen (Hill et al., 1958), and it was conflicting with our result. Feed intake tended to increase in conditions of increasing plasma estrogen concentrations, which was the result of increase in carbohydrate-precursors for lipid synthesis (Hansen and Walzem, 1993). In the present study, we did not measure the levels of plasma estrogen and further research are required to clarify the relationship between plasma estrogen concentration and feed intake in chicks treated with estrogen.
T5 and T6 challenged with estrogen showed significantly higher relative weight of liver than those of others (p<0.05). No other treatment groups significantly differ from each other. The groups consuming high energy diet (T2, T4, and T6) had significantly greater relative weights of abdominal fat than groups consuming low energy diet (T1, T3, and T5). The relative weights of abdominal fat in T5 and T6 treated with estrogen tended to be increased compared to respective energy group.
Estrogen treatment has been found to increase fat content in the liver of male chicks (Courtney et al., 1988) and immature male turkeys (Dashti et al., 1983). High energy consumption was also shown to increase the relative weights of abdominal fat (Deaton and Lott, 1985; Jones and Wiseman, 1985). A previous study demonstrated that a high energy diet increased the relative weights of abdominal fat compared to a low energy diet (Deaton et al., 1983), which is in agreement with the results of the present study.
Serum lipid fractions and enzyme activities
The effects on concentration of lipid fractions and enzyme activities in serum are shown in Table 4. T5 and T6 treated with estrogen showed significantly higher concentrations of serum cholesterol, triacylglycerol and phospholipid than the others (p<0.05). In particular, the serum triacylglycerol concentration was increased 4-5-fold compared to the groups without estradiol benzoate treatment. No other treatment groups showed significant differences between each other.

In hens treated with estradiol benzoate, serum lipid fractions were correlated with increased fatty acid and triacylglycerol biosynthesis in the liver (Hermier et al., 1996). In chicks treated with estrogen, the synthesis and secretion of very low density lipoprotein (VLDL) in the livers rapidly increased within 48 h (Tarlow et al., 1977). Similarly, in male chicks treated with estrogen, increased fatty acid biosynthesis and hyperlipemia were observed after 20 h (Manning et al., 1989). The hepatic lipogenesis, hepatic VLDL assembly and secretion, and plasma VLDL concentration had a close correlation with estrogen administration in Japanese quail hens (Chen et al., 1999). Therefore, the increase in serum lipid fractions of T5 and T6 presumably resulted from changes in hepatic lipogenesis, hepatic VLDL assembly and secretion, and plasma VLDL concentration due to the administration of estrogen.
The levels of serum GOT and GTP are a proper indicator to assay the liver and tissue damages (Lumeij, 1997), and a recent study demonstrated that GOT level was significantly increased in laying hens with FLHS (So et al., 2009). In the present study, there were no significant differences in serum GOT and GPT according to dietary levels of energy and lipotropic factors and estradiol benzoate treatment. It may be assumed that dietary and estradiol treatments did not lead to severe liver damage in growing male chicks.
Contents of hepatic lipid fractions
The effects of dietary energy, lipotropic factors, and estradiol benzoate treatment on the levels of various lipid fractions and MDA in liver are shown in Table 5. The level of hepatic triacylglycerol was increased by more than 10-fold in groups treated with estradiol benzoate, although there was no difference in level of hepatic cholesterol. On the other hand, the level of hepatic phospholipid in chicks treated with estradiol benzoate was significantly decreased as compared with other groups (p<0.05). The contents of various lipid fractions were not influenced by dietary energy level and lipotropic factors. Hepatic MDA levels did not show significant difference among the groups.
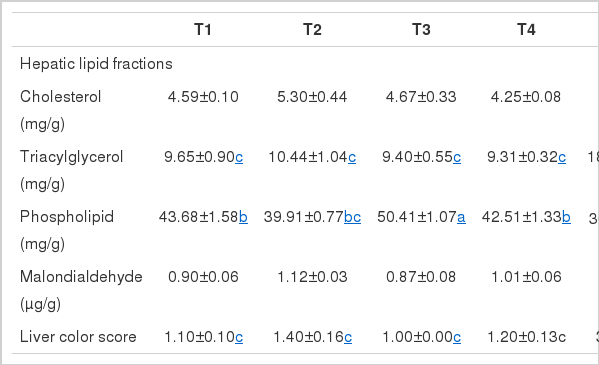
Hepatic fat concentration was linked to quantity of fatty acid synthesis and hepatic lipoprotein. In estrogen-injectd male chicks, the hepatic fat concentration increased 6-fold, likely due to the rise in triacylglycerol levels (Hermier et al., 1996), which is in agreement with the Dashti et al. (1983). In vitro, fatty acid synthesis noticeably increased in chicks implanting 17-β estradiol (Rosebrough et al., 1989). Considering the prior experiment, the acceleration of fatty acid and triacylglycerol synthesis in estrogenized hens in the present study may be a result of increase of hepatic triacylglycerol content. Indeed, intramuscular 17-β estradiol administration induced fatty acid synthesis and triacylglycerol synthesis, and increased enzyme activity (Courtney et al., 1988).
The liver color scores in groups non-treated with estrogen were no difference. However, in groups treated with estrogen, the liver turned ugly yellow, indicated by an increased liver color score. This color change was corresponded with increased hepatic lipid content. In contrast, neither hepatic lipid concentration nor liver color score was affected by dietary energy level or lipotropic factors. This may have resulted from the short experimental period of six weeks, only slight differences in energy levels between the diets or a lack of lipotropic factor deficiency. Nevertheless, these results indicate that estrogenizing is more effective than dietary energy or nutrient level when the young chicks were used as a study model causing FLS and FLHS.
Effects on mRNA related to lipid metabolism
Figure 2 summarizes the effects of dietary components and estrogenization on hepatic mRNA expression related to lipid metabolism as measured by RT-PCR.
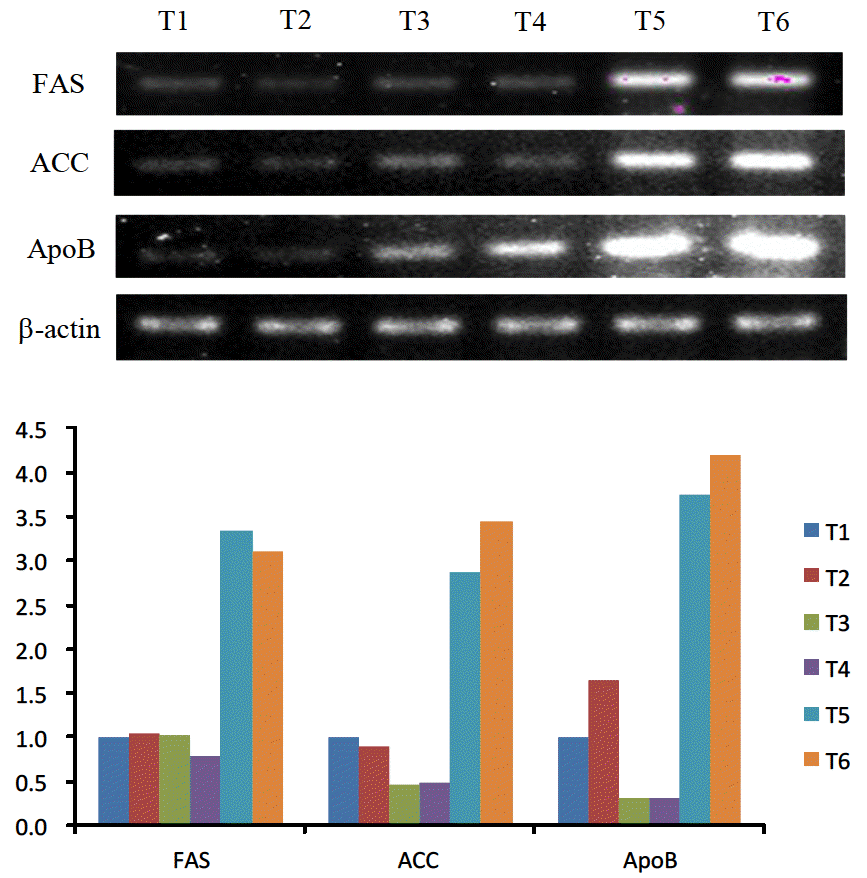
Hepatic mRNA expression. Semi-quantitative RT-PCR analysis. RT-PCR was performed on 1 μg of total RNA using fatty acid synthase, acetyl CoA carboxylase, apolipoprotein B and β-actin specific primers as a control for normalization. The relative levels of fatty acid synthetase, acetyl CoA carboxylase and apolipoprotein B expression were determined by normalisation with respect to β-actin expression. T1 = 11.72 MJ/kg diet+low level lipotropic factors; T2 = 12.89 MJ/kg diet+low level lipotropic factors; T3 = 11.72 MJ/kg diet+high level lipotropic factors; T4 = 12.89 MJ/kg diet+high level lipotropic factors; T5 = 11.72 MJ/kg diet+high level lipotropic factors+2 mg/kg BW estradiol benzoate; T6 = 12.89 MJ/kg diet+high level lipotropic factors+2 mg/kg BW estradiol benzoate.
Lipogenic enzyme genes such as FAS, ACC and ApoB were not affected by the dietary energy level. T5 and T6 which were treated with estrogen expressed 3 times more FAS, 6 to 7 times more ACC, and 12 to 13 times more ApoB than T3 and T4 which were not treated with estrogen but were fed an equivalent level of lipotropic factors as T5 and T6.
Activities and accumulations of ACC and FAS mRNA were indicators to measure the fatty acid metabolism, and these enzymes were affected by nutrition and hormones (Girard et al., 1994; Wolf, 1996). In ovariectomized and estrogen-implanted hens, there was increased activity of these enzymes as well as fatty acid synthesis (Balnave et al., 1978). Consistent with the results of the present study, activities of ACC and FAS mRNA were increased in estrogen-injected male chicks (Manning et al., 1989). Similarly, lipid metabolism was elevated in hens intramuscularly treated with 17-β-estrogen (Courtney et al., 1988).
In this study, T3 and T4 which were treated with high levels of lipotropic factors displayed reduced ACC and ApoB mRNA expression than T1 and T2 which were treated with low levels of lipotropic factors. Specifically, ACC expression was reduced to 1/2 and ApoB expression reduced to 1/5 to 1/3. However, there were no statistically significant difference in concentration of plasma and hepatic lipid fraction. The lack of effect of dietary lipotropic factors on liver and serum lipid fractions was somewhat unexpected. Because available information on the levels of lipid fractions and related enzymes is very few, the interpretation of present result has remained difficult. Among the groups treated with estrogen (T5 and T6), T6 consuming a high energy diet had higher expression of ACC and ApoB mRNA than T5 consuming a low energy diet. However, T5 had higher expression of FAS mRNA than T6, indicating that energy level of diet was not related to mRNA expression.
The average relative weights of liver in the estrogen treated groups were significantly higher than those or non-treated groups, whereas average abdominal fat weights in the hight energy groups were increased as compared to those of the other groups. The treatment of estrogen resulted in significant increase in the concentrations of serum cholesterol, triacylglycerol and phospholipid (p<0.05). The hepatic triacylglycerol contents of estrogenized chicks. There were no significant differences in malondialdehyde concentration among the groups. The estrogen stimulation resulted in dramatic increase in the levels of fatty acid synthetase, acetyl-CoA carboxylase and ApoB mRNA.
ACKNOWLEDGEMENT
The authors acknowledge the financial support of Konkuk University and Cargill Agri Purina, Inc. for the present study.