Influence of Yeast Fermented Cassava Chip Protein (YEFECAP) and Roughage to Concentrate Ratio on Ruminal Fermentation and Microorganisms Using In vitro Gas Production Technique
Article information
Abstract
The objective of this study was to determine the effects of protein sources and roughage (R) to concentrate (C) ratio on in vitro fermentation parameters using a gas production technique. The experimental design was a 2×5 factorial arrangement in a completely randomized design (CRD). Factor A was 2 levels of protein sources yeast fermented cassava chip protein (YEFECAP) and soybean meal (SBM) and factor B was 5 levels of roughage to concentrate (R:C) ratio at 80:20, 60:40, 40:60, 20:80, and 0:100, respectively. Rice straw was used as a roughage source. It was found that gas production from the insoluble fraction (b) of YEFECAP supplemented group was significantly higher (p<0.05) than those in SBM supplemented group. Moreover, the intercept value (a), gas production from the insoluble fraction (b), gas production rate constants for the insoluble fraction (c), potential extent of gas production (a+b) and cumulative gas production at 96 h were influenced (p<0.01) by R:C ratio. In addition, protein source had no effect (p>0.05) on ether in vitro digestibility of dry matter (IVDMD) and organic (IVOMD) while R:C ratio affected the IVDMD and IVOMD (p<0.01). Moreover, YEFECAP supplanted group showed a significantly increased (p<0.05) total VFA and C3 while C2, C2:C3 and CH4 production were decreased when compared with SBM supplemented group. In addition, a decreasing R:C ratio had a significant effect (p<0.05) on increasing total VFA, C3 and NH3-N, but decreasing the C2, C2:C3 and CH4 production (p<0.01). Furthermore, total bacteria, Fibrobacter succinogenes, Ruminococcus flavefaciens and Ruminococcus albus populations in YEFECAP supplemented group were significantly higher (p<0.05) than those in the SBM supplemented group while fungal zoospores, methanogens and protozoal population remained unchanged (p>0.05) as compared between the two sources of protein. Moreover, fungal zoospores and total bacteria population were significantly increased (p<0.01) while, F. succinogenes, R. flavefaciens, R. albus, methanogens and protozoal population were decreased (p<0.01) with decreasing R:C ratio. In conclusion, YEFECAP has a potential for use as a protein source for improving rumen fermentation efficiency in ruminants.
INTRODUCTION
The security of livestock feed is been becoming critical in terms of both quantity and quality, particularly the protein sources, lack of which results in low productivity. Researchers have been trying to find alternative protein sources which could help to increase livestock productivity and efficiency (Wanapat and Rowlinson, 2007; Guglielmelli et al., 2010). Cassava (Manihot esculenta, Crantz) is widely grown in the tropical region (Wanapat, 2003), its’ tuber is a good source of carbohydrate and the leaf as a protein supplement in ruminants (Devendra, 1988; Wanapat, 2003). Cassava root is an excellent source of dietary energy in beef cattle diets (Wanapat and Khampa, 2007). However, it has a readily fermentable carbohydrate with a crude protein level that is low for ruminants (Wanapat, 2003). The process of protein enrichment of animal feed using microorganisms in a semi-solid culture to improve the nutritional value of ruminants feed has been evaluated (Oboh and Akindahinsi, 2003; Oboh, 2006; Aro, 2008). Incorporation of microbial additives such as a culture of Saccharomyces cerevisiae to the diet has become common practice in ruminant nutrition (Campanile et al., 2008). Boonnop et al. (2009) showed that using fermented cassava chip with yeast increased crude protein from 2 to 30.4% CP. Recently, Polyorach et al. (2012; 2013) reported that yeast fermented cassava chip protein (YEFECAP) could be prepared to increase crude protein level up to 47% CP. The beneficial use of YEFECAP has been investigated (Boonnop et al., 2010; Polyorach et al., 2010; Wanapat et al., 2011).
Roughage and concentrate type and ratio in complete feeds is of major importance for efficient utilization of dietary nutrients for production. Moderate amounts of concentrate in the diet can improve the utilization of fiber due to the better supply of fermentable organic matter, energy and nitrogen to rumen bacteria. Earlier work has shown that dietary protein degradation is increased by the inclusion of roughage in the diet (Schoeman et al., 1972), others have reported that protein degradation was either increased by cereal inclusion in the basal diet (Lindberg, 198l) or was unaffected by basal diet (Siddons and Paradine, 1981). Bach et al. (2005) indicated that the most important factors affecting utilization of dietary protein in the rumen included type of protein, carbohydrate and their interactions and the predominant microbial population in the rumen.
However, study of the use of YEFECAP as a protein source with various levels of roughage to concentrate (R:C) ratio have not yet been evaluated. Therefore, the objective of this study was to determine the affect of protein sources with various R:C ratios on ruminal fermentation using an in vitro gas production technique.
MATERIALS AND METHODS
Preparation of yeast-fermented cassava chip (YEFECAP)
YEFECAP preparation was done according to the method of Polyorach et al. (2013) and some important details are as follows: activated yeast was prepared using 20 g of Bakers’ yeast and 20 g cane sugar mixed with 100 mL distilled water, then mixed well and incubated at room temperature for 1 h (A). Liquid media was prepared using 8 g molasses and 100 mL distilled water, followed by addition of 64 g urea, then adjusting the pH of the solution using H2SO4 to achieve a final pH 3.5 to 5 (B). Mixed (A) and (B) at 1:1 ratio then flushed with air for 66 h at room temperature using an air pump (600 W). After 66 h, the yeast medium solution twas mixed with cassava chips at a ratio of 1 mL: 1.3 g, then fermented in solid state under shade for 72 h, followed by sun-drying for 48 h. The final product is stored in plastic bag for later use as an ingredient in the concentrate supplement.
Experimental design and dietary treatments
This study was conducted using an in vitro gas production technique at various incubation time intervals. The experimental design was a 2×5 factorial arrangement in a completely randomized design (CRD) with three replications per treatment. The treatments were two sources of protein in concentrate in which concentrate I used YEFECAP and concentrate II used soybean meal (SBM) as protein sources with five levels of R:C ratio at 80:20, 60:40, 40:60, 20:80, and 0:100, respectively. Rice straw was used as a roughage source. Samples of roughage and concentrates were dried at 60 °C, then ground to pass a 1-mm sieve (Cyclotech Mill, Tecator, Sweden) and used for chemical analysis and in the in vitro gas test. The samples were analyzed for dry matter (DM), ash and crude protein (CP) using the procedures of AOAC (1998), neutral detergent fiber (NDF) and acid detergent fiber (ADF) according to Van Soest et al. (1991).
Animals and preparation of rumen inoculums
Two, male 2-year-old, rumen fistulated dairy steers with an initial BW of 280±15.0 kg were used as rumen fluid donors. Dairy steer rumen fluid was collected from animals fed with concentrate (18.0% CP and 80.6% TDN) at 0.5% of BW in to equal portions, at 07.00 h and 16.00 h and rice straw was fed on ad libitum basis. The animals were kept in individual pens and clean fresh water and mineral blocks were offered as free choice. The animals received the diets for 20 d before the rumen fluid was collected. On d 20, 1,000 mL rumen liquor was obtained from each animal before the morning feeding. The rumen fluid was filtered through four layers of cheesecloth into pre-warmed thermo flasks and then transported to the laboratory.
In vitro fermentation of substrates
Samples of each total mixed substrate (200 mg), following respective treatments were weighed into 50 mL serum bottles. For each treatment, three replications were prepared. Ruminal fluid from each animal was mixed with the artificial saliva solution of Menke and Steingass (1988) in a proportion 2:1 (mL/mL) at 39°C under continuous flushing with CO2. Thirty milliliters of rumen inoculum mixture were added into each bottle under CO2 flushing. Bottles were sealed with rubber stoppers and aluminium caps and incubated at 39°C (96 h) for in vitro gas test. Thirty minutes after starting the incubation, the bottles were gently mixed and then mixed three times every 3 h. For each sampling time, three bottles containing only the rumen inocula were included within each run and the mean gas production values of these bottles were used as blanks. The blank values were subtracted from each measured value to give the net gas production.
Sample and analysis
During the incubation, data of gas production was measured immediately after incubation at 0, 2, 4, 6, 8, 12, 18, 24, 48, 72, and 96 h by using a pressure transducer and a calibrated syringe. Cumulative gas production data were fitted to the model of Orskov and McDonald (1979) as follows:
Where a = the gas production from the immediately soluble fraction, b = the gas production from the insoluble fraction, c = the gas production rate constant for the insoluble fraction (b), t = incubation time, (a+b) = the potential extent of gas production. y = gas produced at time “t”.
Inoculum ruminal fluid was sampled at 0, 4, 6, 12, and 24 h post inoculations. Rumen fluid samples were then filtered through four layers of cheesecloth. Samples were divided into 3 portions; the first portion was centrifuged at 16,000×g for 15 min, and the supernatant was stored at −20°C before NH3-N analysis using the micro-Kjeldahl methods (AOAC, 1998) and VFA analysis using HPLC (Samuel et al., 1997). The second portion was fixed with 10% formalin solution in a sterilized 0.9% saline solution for a total direct count of bacteria, protozoa and fungi made by the methods of Galyean (1989) based on the use of a hemocytometer (Boeco, Hamburg, Germany). The final portion was stored at −20°C for DNA extraction (Yu and Morrison, 2004).
In vitro degradability was determined after termination of incubation, when the contents were filtered through pre-weighed Gooch crucibles and residual dry matter was estimated. The percent loss in weight was determined and presented as in vitro dry matter degradability (IVDMD). The dried feed sample and residue left from above was ashed at 550°C for determination of in vitro organic matter degradability (IVOMD) (Tilley and Terry, 1963). Calculation of ruminal methane (CH4) production using VFA proportions was made according to Moss et al. (2000) and as follows:
Rumen microbial population
Community deoxyribonucleic acids (DNA) extraction
Community DNA was extracted from 0.5 g of rumen content (fluid and digesta) by the RBB+C method (Yu and Morrison, 2004). In brief, the RBB+C method employs two rounds of bead beating in the presence of NaCl and SDS, followed by sequential ammonium acetate and isopropanal precipitations. The precipitated nucleic acids were then treated with RNase A and proteinase K, and the DNA was purified using columns from QIAgen DNA Mini Stool Kit (QIAGEN, Valencia, CA).
Primers and polymerase chain reaction (PCR)
The targeted bacteria were total bacteria, the three predominant cellulolytic bacteria (Fibrobactor succinogenes, Ruminococcus flavefaciens and Ruminococcus albus), methanogens and protozoa. Primers for F. succinogenes, Fs219f (5′GGT ATG GGA TGA GCT TGC-3′) and Fs654r (5′-GCC TGC CCC TGA ACT ATC- 3′), were selected to allow amplification (446-bp product) of all 10 F. succinogenes strains deposited in GenBank. For R. albus primers, Ra1281f (5′-CCC TAA AAG CAG TCT TAG TTC G-3′) and Ra1439r (5′ CCT CCT TGC GGT TAG AAC A- 3′) (175-bp product). R. flavefaciens primers, Rf154f (5′-TCT GGA AAC GGA TGG TA-3′) and Rf425r (5′-CCT TTA AGA CAG GAG TTT ACA A-3′), were also selected to allow species–specific amplification (295 bp) of all seven R. flavefaciens strains deposited in GenBank. All these primer sets were previously published by Koike and Kobayashi (2001). For methanogens primers, forward primers (5′-TTCGGTGGATCDCARAGRGC-3′) and reverse primers (5′-GBARGTCGW- AWCCGTAGAATCC -3′) were employed as described by Denman et al. (2005). PCR conditions for F. succinogenes were as follows: 30 s at 94°C for denaturing, 30 s at 60°C for annealing and 30 s at 72°C for extension (48 cycles), except for 9 min denaturation in the first cycle and 10 min extension in the last cycle. Amplification of 16S rRNA for the other two species was carried out similarly except an annealing temperature of 55°C was used. The PCR conditions for methanogens were as follows; 30 s at 94°C for denaturing, 30 s at 58°C for annealing and 90 s at 72°C for extension (35 cycles) (Wright et al., 2004) and ruminal protozoa was described by Sylvester et al. (2004).
Real-time PCR
The targeted bacteria were cellulolytic bacteria (F. succinogenes, R. albus, and R. flavefaciens) and methanogens. To establish a quantitative assay, we amplified target 16s rDNA of each species by using specific primers and PCR conditions as described previously, the purified DNA was quantified by spectrophotometry by comparing the products with serial 10- fold dilutions from 101 to 108 DNA copies of the previously quantified DNA standards. Real-time PCR amplification and detection were performed in a Chromo 4TM system (Bio-Rad, USA). In brief, Biostools QuantiMix Easy SYG Kit was used for PCR amplification. Samples were assayed in duplicate in a 20 μL reaction mixture contained 4 to 6 mM MgCl2, 10 μL of Mastermix (including; Taq DNA polymerase, reaction buffer, dNTP mixture, MgCl2 and SybrGreen), 2 μL of DNA template, and 0.8 μL of each primer (10 μM/μL).
Statistical analysis
All data were analyzed as a 2×5 factorial arrangement in a Completely randomized design (CRD) using the PROC GLM of SAS (1998). Data were analyzed using the model:
Where: Y = observations; μ = overall mean; Ai = effect of factor A (protein sources, i = 1 to 2); Bj = effect of factor B (level of roughage to concentrate (R:C) ratio, j = 1 to 5), ABij = interaction between factor A and B, and ɛij = the residual effect. Multiple comparisons among treatment means were performed by Duncan’s New Multiple Range Test (DMRT) (Steel and Torrie, 1980). Differences among means with p<0.05 were accepted as representing statistically significant differences. Trend of R:C ratio level was performed by orthogonal polynomials comparison.
RESULTS
Feed ingredients and chemical compositions of concentrate I, concentrate II, YEFECAP and rice straw are presented in Table 1. Concentrate I, using YEFECAP while concentrate II using SBM as main protein sources. Chemical composition of both concentrate I and concentrate II were similar between treatments. YEFECAP product contained DM, OM, CP, NDF, and ADF at 89.2, 95.6, 46.7, 6.7, and 4.2% DM, respectively. Rice straw was used as a roughage source which contained DM, OM, CP, NDF, and ADF at 90.2, 81.3, 2.9, 82.7, and 57.6% DM, respectively.
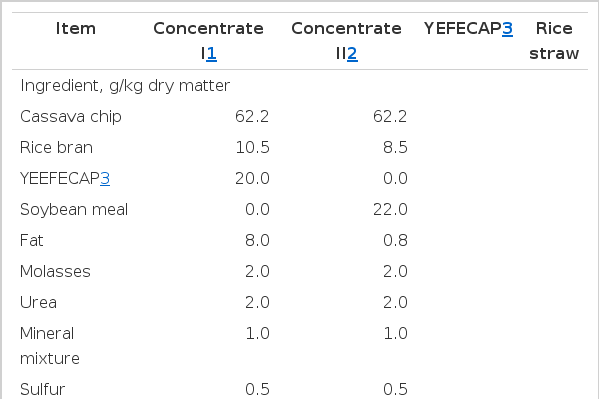
Feed ingredients composition of dietary treatments used in the experiment
Cumulative gas production for each of the substrate treatments is presented as gas production and the values for estimated parameters obtained from the kinetics of gas production models for substrates studied are given in Table 2. Protein source had no effect (p>0.05) on the kinetics of gas production except the gas production in the insoluble fraction (b) of YEFECAP group was significantly higher (p<0.05) than in the SBM group. Moreover, the intercept value (a), gas production from the insoluble fraction (b), gas production rate constants for the insoluble fraction (c), potential extent of gas production (a+b) and cumulative gas production at 96 h were influenced (p<0.01) by the R:C ratio.
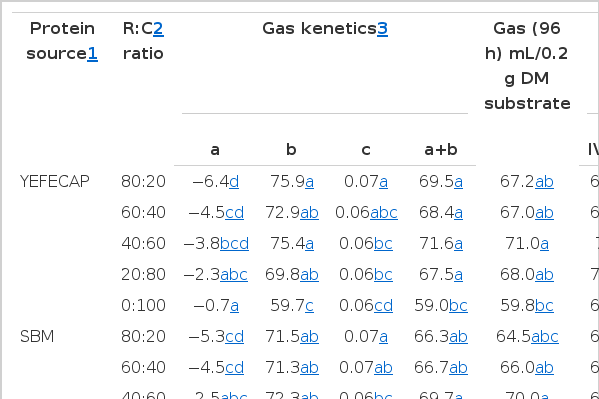
Effect of protein source with roughage to concentrate ratio on gas production kinetics and degradability from in vitro incubation with rumen fluid
In addition, the effect of protein source and R:C ratio on in vitro degradability is presented in Table 2. Protein source had no effect (p>0.05) on either IVDMD or IVOMD while R:C ratio increased (p<0.01) IVDMD and IVOMD when the R:C ratio level was decreasing and were highest in R:C ratio at 40:60. However, there were interactions between protein sources and R:C ratio on gas production kinetics and degradability.
The VFA, NH3-N and methane production (CH4) are presented in Table 3. The results of the current study revealed that the YEFECAP fed group significantly increased (p<0.01) total VFA and propionate (C3) while decreased (p<0.05) acetate to propionate ratio (C2:C3) and CH4 production when compared with SBM fed group. However, with a decreasing R:C ratio level, total VFA, C3 and NH3-N were increased (p<0.05) while C2, C2:C3 and CH4 production were decreased (p<0.05). Moreover, there were interactions between protein sources and R:C ratio which affected (p<0.05) on C2, C3 and C2:C3.
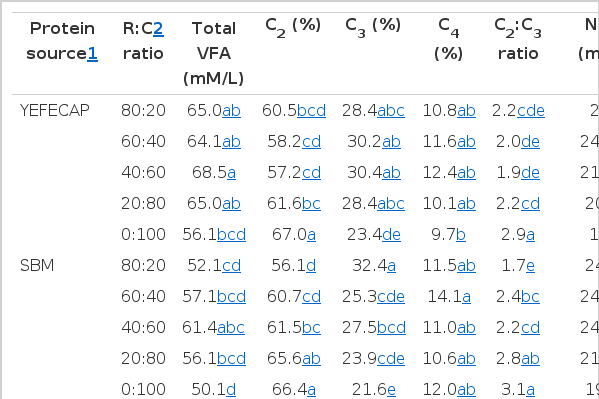
Effect of protein source with roughage to concentrate ratio on in vitro volatile fatty acids (VFA), ammonia nitrogen (NH3-N) and methane production (CH4)
Effect of protein source and R:C ratio on ruminal microorganisms is presented in Table 4. The results revealed that the bacterial population in YEFECAP fed group were significantly higher (p<0.01) than in the SBM group while fungal zoospores and protozoal population were not significantly different (p>0.05) between the two sources of protein. However, with a decreasing R:C ratio level, bacterial and fungal zoospores populations were increased (p<0.01) while the protozoal population was decreased (p<0.01). External standards for real-time PCR were prepared from a simulated rumen matrix. For each standard, linear regressions derived from the threshold cycle (C[T]) of each DNA dilution versus the log quality were calculated. Logarithms of the DNA concentration (copies/mL) were plotted against the calculated means, obtaining straight line equations y = −0.3541x+11.29, y = −0.3094x+12.48, y = −0.3766x+10.38, y = −0.3293x+11.03, y = −0.3279x+11.23 and y = −0.2176x+10.66 (where y is the log of DNA concentration and x is the Ct), with a linear correlation coefficient (r2) of 0.986, 0.985, 0.996, 0.993, 0.998, and 0.992 for total bacteria, F. succinogenes, R. flavefaciens, R. albus, methanogens and protozoa, respectively. The accuracy of each real-time PCR was validated by quantifying known numbers of target species templates (total bacteria, F. succinogenes, R. flavefaciens, R. albus, methanogens and protozoa) and is shown in Table 4. It was shown that the interaction between protein source and R:C ratio did not affect populations of protozoa, total bacteria, methanogens and predominant cellulolytic bacteria except R. flavefacieus. Whereas, total bacteria, F. succinogenes, R. flavefacieus, and R. albus in the YEFECAP supplemented group were significantly higher (p<0.05) than in the SBM supplemented group. Moreover, when increasing concentrate level, total bacterial and fungal zoospores population were increased (p<0.01) and the highest in R:C ratio was found at 40:60, while, F. succinogenes, R. flavefaciens, R. albus, protozoal population and methanogens were significantly decreased (p<0.01) with increasing concentrate level.
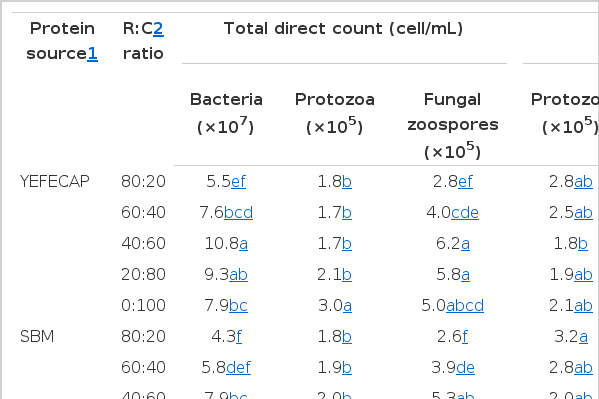
Effect of protein source with roughage to concentrate ratio on ruminal microorganism
DISCUSSION
Under this experiment, the rice straw that was used as a main roughage source contained 2.9% CP (Table 1). This value was similar to those values reported by Wanapat et al. (2009). Moreover, crude protein of YEFECAP was similar to the value reported earlier by Polyorach et al. (2012; 2013) while this value was relatively higher than those reported by Boonnop et al. (2009) and Wanapat et al. (2011), which could be due to the differences of proportion of urea and molasses used in the liquid medium, as well as the proportions of yeast fermented liquid and cassava chips used.
The present result revealed that gas production in the insoluble fraction (b) in YEFECAP supplemented group was significantly higher (p<0.05) than in SBM supplemented group. This was probably due to YEFECAP promoting growth of rumen cellulolytic bacteria and lactate-utilizing bacteria. Yeast may also have stimulated bacterial growth through removal of oxygen that occurs in ruminal fluid and in that way can prevent toxicity to the ruminal anaerobes (Chaucheyras-Durand et al., 2008; Doto and Liu, 2011). This will in turn increase polysaccharidase and glycoside-hydrolase activities towards lignified plant tissues, “unlocking” the fiber’s digestible fraction (hemicellulose and cellulose), which will be digested by the cellulolytic microorganisms. These results were similar to the finding of Wanapat et al. (2011) that YEFECAP can fully replace SBM in concentrate for dairy cows and improved rumen fermentation, dry matter intake, nutrient digestibility, milk production and composition, as well as the improving the economic return in early lactating dairy crossbreds.
Gas kinetics and cumulative gas production were influenced by different R:C ratios, especially the constant rate (c) and in vitro degradability of both DM and OM which were increased (p<0.01) with an increasing level of concentrate and the highest was in R:C at 40:60. These could be due to increased concentrate level that would provide more readily available energy, enhancing corresponding of microbes, consequently, increased degradability. The stimulatory effect of concentrate feeds on the ruminal microflora is well documented and, according to Hungate (1966) who reported that a pronounced effect in the rumen wamore readily achieved from carbohydrate than from forages. These findings were in agreement with Lunsin and Wanapat (2010) and Anantasook and Wanapat (2012). Moreover, Cherdthong et al. (2010) reported that the value for NDF and ADF were decreased particularly with the highest level of concentrate. However, a high level of concentrate may lower the pH and impact on the microbial population (Slyter, 1976).
The level of NH3-N concentration increased with an increasing concentrate level and the values ranged from 19.8 to 26.6 mg/dL which were similar to the values reported by Wanapat and Pimpa (1999) (15 to 30 mg/dL in the rumen).
In addition, YEFECAP as a protein source group significantly increased (p<0.05) total VFA and C3 while decreased (p<0.05) C2:C3. These results suggested that YEFECAP is a good source of protein as Wanapat et al. (2011) also reported that total VFA and C3 production were significantly increased, consequently, C2:C3 ratio was reduced in the 100% YEFECAP supplemented group. Moreover, this was probably due to some effects of the yeast contained in YEFECAP that stimulated the growth and metabolism of rumen microorganisms especially lactate-utilizing bacteria, such as Megasphaera elsdenii or Selenomonas ruminantium (Lynch and Martin, 2002). In addition, yeast could supply different growth factors, such as amino acids, peptides, vitamins, and organic acids, essential for the ruminal bacterial growth (Chaucheyras-Durand et al., 2008), hence enhancing VFA concentration and reducing C2:C3 proportion (Oeztuerk, 2009).
Furthermore, total VFA and C3 were increased while C2, and C2:C3 ratio were decreased with a increasing concentrate level. Similar to the current experiment, an enhanced propionate concentration was found in the rumen of cows fed high-grain as compared with high-forage diets (Sutton et al., 2003; Calabrò et al., 2008; Zicarelli et al., 2011). Surprisingly, Moorby et al. (2006) reported linear increases in total VFA and butyrate concentrations and a decrease in acetate with increasing proportion of concentrate in dietary DM, but the concentration of propionate was not affected. Calsamiglia et al. (2008) and Cherdthong et al. (2010) also reported that high concentrate diet fermentation resulted in a greater molar proportion of ruminal propionate. The inverse of relationship between C2:C3 ratio and amount of concentrate in the diet has often been explained by the tendency of fiber fermenting bacteria to produce C2 and starch fermenting bacteria to produce C3 (Slyter, 1976).
CH4 production in the rumen of YEFECAP supplemented group was lower (p<0.05) when compared with SBM group. This could be due to the yeast in YEFECAP, probably affecting hydrogen metabolism in the rumen. Yeast also has the potential to alter the fermentation process in the rumen in a manner that reduces the formation of methane (CH4) gas. Previously, Lynch and Martin (2002) reported a 20 mM/L reduction of methane production after 48 h of incubation of mixed rumen microorganisms in the presence of alfalfa and a live yeast product. McGinn et al. (2004) showed a 3% (g/kg of DMI) decrease in in vivo CH4 production for one commercial yeast product. Moreover, CH4 production was also influenced by R:C ratio, when an increasing concentrate level decreased (p<0.01) methane production and the lowest value was at 0:100. This result might be due to an increased proportion of concentrate in the diet which changes ruminal VFAs concentrations in such a way that less C2 and more C3 is formed, and hence, the supply of hydrogen for methanogenesis is limited. In addition, concentrate feeding has been shown to reduce methane output by reducing the protozoal population (Van Soest, 1982; Iqbal et al., 2008). The amount of CH4 produced varies according with the type of diet (forage/concentrate) and the type of production system (intensive/extensive) (Moss et al., 2000; Aluwong et al., 2011). A positive response to high levels of starch-based concentrate (grains) on methane reduction has also been reported by others (Yan et al., 2000; Beauchemin and McGinn, 2005; Lovett et al., 2005; Poungchompu et al., 2009).
The results showed that the bacterial population in YEFECAP supplemented group was significantly higher (p<0.01) than in the SBM supplented group while fungal zoospores and protozoal population were not changed (p>0.05). These results related with the data from real-time PCR technique, regarding total bacteria, F. succinogenes, R. flavefacieus, and R. albus in the YEFECAP fed group which were significantly higher (p<0.05) than in the SBM fed group which received a good protein source and the presence of yeast. Jouany (2006) reported that there were three main roles of yeast in the rumen that have been identified: i) Yeasts are source of nutrients for rumen microbes, ii) Yeasts could consume oxygen which is present in the microenvironment surrounding solid particles which have just been ingested into the rumen, iii) S. cerevisiae was able to out compete S. bovis for the utilization of sugars. These attributes create an optimal environment for rumen microbe activity especially cellulolytic bacteria. Additionally, S. cerevisiae leads to increased germination of zoospores from a rumen fungal strain of Neocallimastix frontalis as shown in an in vitro study (Chaucheyras et al., 1995). Callaway and Martin (1997) also showed that cellulolytic bacteria became established earlier, and remained at a high and stable level, even after a particularly stressful period when lambs, fitted with a rumen cannula, were fed an active dry yeast (ADY) daily. The increasing of cellulolytic bacteria was also mesured with real-time PCR as reported by Mosoni et al. (2007) who showed that proportions of 16S ribosomal RNA of the three main cellulolytic bacterial species (F. succinogenes, R. albus, and R. flavefaciens) increased in the rumen of sheep fed with yeast.
These results agreed with Boonnop et al. (2010) who found that YEFECAP could completely replace soybean meal and was beneficial to cattle in terms of efficiency of rumen fermentation, microbial protein synthesis, nitrogen retention and nutrient digestibilities. Polyorach et al. (2010) and Wanapat et al. (2011) later reported that increasing the level of YEFECAP in concentrate mixtures linearly increased the population of bacterial and fungal zoospores in the rumen while protozoa were not changed.
In addition, populations of fungal zoospores and total bacteria were significantly increased (p<0.01) with the highest levels found when the R:C ratio was at 40:60, while predominant cellulolytic bacteria (F. succinogenes, R. flavefaciens, and R. albus), methanogens and protozoal population were decreased with a decreasing level in the R:C ratio. Decreasing of the three main cellulolytic bacterial populations in this study might be due to concentrate-rich diets. The influence of concentrate on fiber digestion and roughage utilization depends also on composition and ruminal fermentability of concentrate feed in the diet (Choi et al., 2003; Tafaj et al., 2005). Similarly to Cherdthong et al. (2010) who reported that R:C ratio of 100:0, 75:25, 50:50, and 25:75 could increase bacterial and fungal population while decreased numbers of the three main cellulolytic bacteria (F. succinogenes, R. flavefaciens and R. albus) occurred with increasing concentrate levels. Waraanu et al. (2000) reported that roughage- to-concentrate ratios of 100:0, 60:40, and 40:60 could decrease the cellulolytic bacterial population in swamp buffalo (5.62×1010, 4.06×1010, and 4.57×1010 CFU/mL), respectively. Moreover, Anantasook and Wanapat (2012), using the in vitro gas production technique, also confirmed that the bacterial population was increased while cellulolytic bacterial and protozoal populations were decreased with increasing concentrate level.
CONCLUSIONS
Based on this study, it could be concluded that using YEFECAP as a protein source with R:C ratio of 40:60 could improve in vitro nutrient degradability and rumen fermentation by increasing total VFA, C3, fungal zoospores and total bacteria while C2, C2:C3 ratio, CH4 production, methanogens and protozoal population were decreased. These results reveal a potential use of YEFECAP as a protein source, leading to improving rumen fermentation efficiency and a possible productivity in ruminants.
ACKNOWLEDGEMENTS
The authors would like to express their most sincere thanks to the Tropical Feed Resources Research and Development Center (TROFREC), Department of Animal Science, Faculty of Agriculture, Khon Kaen University, Khon Kaen, Thailand, The Thailand Research Fund (TRF) through the Royal Golden Jubilee Ph. D. Scholarship Program for providing financial support of research and the use of research facilities.