Dietary Supplementation of Benzoic Acid and Essential Oil Compounds Affects Buffering Capacity of the Feeds, Performance of Turkey Poults and Their Antioxidant Status, pH in the Digestive Tract, Intestinal Microbiota and Morphology
Article information
Abstract
Three trials were conducted to evaluate the effect of supplementation of a basal diet with benzoic acid or thymol or a mixture of essential oil blends (MEO) or a combination of benzoic acid with MEO (BMEO) on growth performance of turkey poults. Control groups were fed a basal diet. In trial 1, benzoic acid was supplied at levels of 300 and 1,000 mg/kg. In trial 2, thymol or the MEO were supplied at levels of 30 mg/kg. In trial 3, the combination of benzoic acid with MEO was evaluated. Benzoic acid, MEO and BMEO improved performance, increased lactic acid bacteria populations and decreased coliform bacteria in the caeca. Thymol, MEO and BMEO improved antioxidant status of turkeys. Benzoic acid and BMEO reduced the buffering capacity compared to control feed and the pH values of the caecal content. Benzoic acid and EOs may be suggested as an effective alternative to AGP in turkeys.
INTRODUCTION
The routine use of in-feed antibiotics to promote growth has been questioned due to the potential development of resistance to a number of pathogenic bacterial species (Wegener et al., 1998). Fear of transmission of this resistance to humans through the food chain has led to precautionary action to exclude several antibiotics from productive animal diets. As a result, in the European Union a complete ban on the use of antibiotic growth promoters has been established since 2006. This ban has been also established in Canada, Korea, and may be enforced in both North and Latin American countries and Australia and is generally acknowledged that all antibacterial feed additives will be banned in the near future (Franz et al., 2010). Therefore, there is a need for intensive research into the identification and evaluation of alternatives to traditional antibiotic feed additives that would satisfy consumer perceptions and would be closer to environmentally friendly farming practices such as organic acids and herb extracts. Although most of these natural approaches have already been used in combination with in-feed antibiotics, their efficacy as the only dietary growth promoting additives has not been yet fully established (Franz et al., 2010).
Benzoic acid and its salts are used as feed additives in fur animals (Polonen et al., 2000) and pigs (Mroz et al., 2000) and in forages for ruminants (Wildgrube and Zausch, 1971). It is well known that benzoate inhibits fungal growth. Today, its antifungal action is widely used (at concentrations of 5 to 10 mM) for the preservation of acid foods such as fruit juices, pickles, wine and pharmaceutical preparations. It is known that antifungal effects are based on the accumulation of benzoate at low external pH, which lowers the intracellular pH into the range where phosphofructokinase is sensitive (Krebs et al., 1983). Probably, a similar mode of action is involved in its antibacterial properties. It has been reported that benzoic acid plays an important role in lowering numbers of many pathogenic bacteria as Campylobacter jejuni, Escherichia coli, Listeria monocytogenes, and Salmonella enterica (Friedman et al., 2003). In previous experiments, bacteriostatic effects of benzoic acid in broiler chicken GIT were demonstrated. However, dosages of this acid higher than 0.25% caused growth depression and poor feed conversion (Józefiak et al., 2006; 2010) while limited information about the effects of benzoic acid on turkey performance is available.
Certain herbs might be an interesting alternative feed supplement to the antibiotic growth promoters. In recent years, many herbal plants such as rosemary, sage, thyme, oregano and tea or their extracts have attracted wide research interest due to antioxidative, antibacterial and antifungal properties (Giannenas et al., 2003; 2005) that are attributed to a great variety of phenolic compounds occurring in these plants. Following their ingestion, these compounds might scavenge the free radicals formed during the metabolic processes in the cells diminishing the oxidative stress (Salah et al., 1995), and improving thus the health status and the performance of productive animals.
Growth performance is a multifactorial dependant. Without the use of antimicrobial drugs, birds’ health, intestinal communities, digestibility and feed cost must be supported by alternative means. However, published information giving evidence that botanical feed additives could efficiently replace the antibiotic growth promoters is yet limited (Franz et al., 2010). The objective of this study was to evaluate the effect of benzoic acid and essential oil compounds on the performance of turkeys.
MATERIAL AND METHODS
Animal’s ethics
The trial protocol was approved by the Institutional Committee of The Veterinary Faculty of the University of Thessaly. Throughout the trial, the turkeys were handled according to the principles for the care of animals in experimentation (National Research Council, 1996).
Animals and diets
A series of three growth trials were performed in a commercial turkey farm, in Basilica (40°28′45″N; 23°08′09″E) Greece. All subgroups were housed in separate floor pens with clean wood shavings, each equipped with an infrared lamp. The lighting regimen provided 24 h of continuous light per day until day 2nd of experiment and 20 h of light per day until day 21st and consecutive cycles of 8 h light and 4 h dark, thereafter. To meet the nutrient requirements of the turkey poults during the experimental period, three complete basal diets were formulated each one for the starter, growing and finishing period, respectively. Feed and drinking water were offered to all birds ad libitum throughout the experiment.
The composition of the basal diet is presented in Table 1. All birds were weighed individually at the time of their placing into the poultry house and later on every week. Feed consumption within each group was recorded weekly and feed conversion ratio was finally calculated. Mortality was also daily recorded.
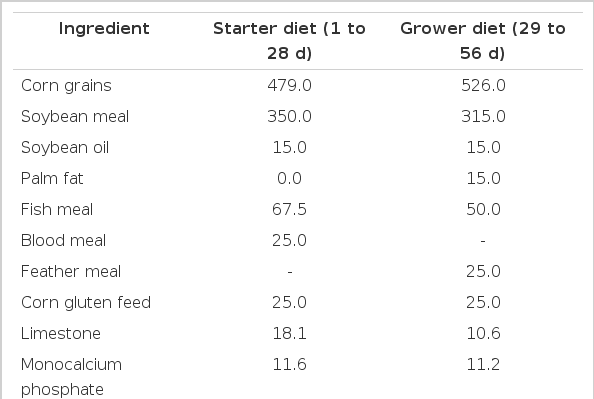
Ingredients and nutrient composition of the basal diet (g/kg)
Trial 1
This trial was conducted with 180-d-old male (Nicholas 300) turkey poults were randomly allocated into three groups of ten birds each with six replicates. During the feeding period that lasted 56 days, one group (control) was fed on a basal diet, the other groups on the same diet supplemented further with either 300 mg/kg benzoic (B300) acid or 1,000 mg/kg (B1,000).
Buffering capacity of the feeds
In order to determine the buffering capacity of the experimental diets and their ingredients using a WTW pH meter. A portion of 10 g feed was placed in a beaker and 100 mL of distilled water were added. The mixture was stirred for about 30 min, and subsequently was titrated with 0.1 N HCl, under continuous stirring, to reach pH 4 (Florou-Paneri et al., 2001). The microliters of the acid consumed were used as the units for expressing the buffering capacity of the feeds.
pH measurements in the digestive tract
At the end of the trial, 3 turkeys from each subgroup were sacrificed by cervical dislocation. The contents of the crop, gizzard, ileum, caeca, and rectum were quantitatively collected. The pH in the contents of all gastrointestinal segments was measured with a combined glass/reference electrode (WTW pH meter, Weilheim, Germany).
Determination of intestinal microbiota
To determine microbial populations, diluted digesta were suspended in pre-reduced salt medium and homogenized for 2 min in CO2-flushed plastic bags using stomacher homogenizer (Interscience, Saint Nom La Bretéche, France). Subsequently, serial decimal dilutions were made, avoiding aeration, using the medium as previously described (Giannenas et al, 2011). Samples incubated under anaerobic conditions at 37°C for 48 h on MRS agar medium (Merck 1.10660, Darmstadt, Germany) were used for the determination of total numbers of lactic acid bacteria, whereas samples incubated under aerobic conditions at 37°C for 24 h on MacConkey agar (Merck 1.05465) were used for the determination of total numbers of coliform bacteria. Results were expressed as base-10 logarithm colony-forming units per gram of ileal or cecal digesta.
Trial 2
In this trial, a total of 180-d-old male (Nicholas 300) turkey poults were randomly allocated into three groups of ten birds each with six replicates. During the feeding period (56 days), one group (control) was fed on the basal diet, while the other groups were fed on the same diet further supplemented with either 30 mg/kg thymol (T30) or 30 mg/kg of a mixture of essential oil compounds (MEO30). The mixture of essential oils was a commercial product (CRINA Poultry-CP; DSM Nutritional Products Ltd, Basel Switzerland) containing a mixture of EO compounds (thymol [≥10%], eugenol [≥0.5%], piperine [≥0.05%] and other flavoring substrates [≤0.6%]).
Antioxidant status determination
Three birds per subgroup were randomly selected for tissue sampling. Following slaughter, liver, breast and thigh muscles tissues were collected by removing skin, fat and connective tissue. Samples were vacuum packaged and stored at −80°C pending further analysis. All excised tissues were assayed for the levels of glutathione peroxidise (GSH-Px), glutathione S-transferase (GST) and malondialdehyde (MDA) formation as described in the following paragraphs. To assess the effect of dietary treatment on lipid oxidation of raw tissue during refrigerated storage, samples were thawed, wrapped in transparent oxygen-permeable polyvinyl chloride film (6,000 to 8,000 cm3/m2×24 h), placed in a non-illuminated refrigerated cabinet at 4°C for 5 days, and submitted to determination of antioxidant enzyme activities and lipid oxidation at 0 and 5 days of refrigerated storage.
Assay of glutathione peroxidase
Activity of GSH-Px was assayed with the method of Paglia and Valentine (1967) with some modifications. Each 1 mL sample contained 25 μL tissue homogenate, 1 mM EDTA, 1 mM sodium azide, 5 units glutathione reductase (GR), 5 mM GSH, 0.2 mM NADPH, and 50 mM phosphate buffer (pH 7.0). The samples were incubated in a 37°C water bath for 30 min and then transferred to cuvettes. A quantity of 17.64 mM H2O2 was added and rapidly mixed and the absorption ratio was measured in a spectrophotometer (Hitachi U-1900, Tokyo, Japan) at 340 nm for 3 min. The absorbance rate (decrease) was calculated based on NADPH consumed using the extinction coefficient for NADPH (0.00622/nmol/mL/cm), corrected to express dilution of NADPH per mL of tissue homogenate expressed as munits/mg protein.
Assay of glutathione S-transferase
Activity of GST was measured by a modified version of Habig et al. (1974). A GST reagent mixture was made, consisting of 50 mL of phosphate buffer (pH 6.5) and 2 mL of 20 mM CDNB (ethanol solution). A quantity of 800 μL of this reagent, along with 100 μL of 5 mM GSH were mixed in a cuvette and subsequently 100 μL of 1:100 the diluted sample were added. The cuvette was immediately inserted into a spectrophotometer and absorbance at 340 nm was read for 3 min. GST activity was expressed as mmol/min/mg protein (extinction coefficient = 9.6).
Lipid peroxidation assay
Malondialdehyde (MDA) was used as a marker of lipid peroxidation using a modified version of the method described by Buege and Aust (1978). Briefly 14 μL of butylated hydroxytoluene and 1,400 μL of a mixture of 0.375 g/L thiobarbituric acid and 0.9 g/L trichloroacetic acid in 0.25 N HCl were added to 100 μL of tissue homogenate (Ultraturrax IKA T18 basic; IKA, Jacqvepagua, Brazil); samples (0.5 g) and placed in tubes. Samples were incubated at 100°C in a water bath for 15 min, centrifuged (Centurion; Scientific Ltd. Company, West Sussex, UK) at 13,000×g for 5 min and the absorbance of the resulted supernatants was read at 532 nm. Concentration of MDA in the samples was plotted against a reference curve made using known amounts of MDA and expressed as nmol/mg of protein. Proteins were determined by the method of Bradford (1976) using bovine serum albumin as a standard.
Further parameters that were evaluated in this trial were measurements of pH in the digestive tract of birds and determination of intestinal microbiota.
Trial 3
A total of 120-d-old male (Nicholas 300) turkey poults were randomly allocated into two groups of ten birds each, with six replicates. During the feeding period that lasted 56 days, one group (control) was fed on a basal diet while the second group was fed on the same diet supplemented with 300 mg/kg of a mixture of benzoic acid and essential oils (BMEO) compounds. The mixture of benzoic acid and essential oil compounds was the commercially available product CRINA Poultry Plus (CPP) containing a mixture of EO compounds: thymol (≥1%), eugenol (≥0.5%), piperine (≥0.05%) and other flavoring substrates (≤0.6%) with benzoic acid (≥80%).
Intestinal morphology measurements
Morphometric analysis of the small intestine was evaluated according to Giannenas et al. (2011). During necropsy of the selected birds, the gastrointestinal tract was removed and the small intestine was divided into three parts: duodenum, jejunum and ileum. One cm-long segments were taken from the center of each part and fixed in 10% buffered formalin for morphometrical assays under light microscopy. Formalin-fixed intestinal tissues were processed, embedded in paraffin wax, sectioned at 3 μm and stained the haematoxylin-eosin method. Histological sections were examined with a Nikon phase contrast microscope coupled with a Microcomp integrated digital imaging analysis system (Nikon Eclipse 80i, Nikon Co., Tokyo, Japan). Images were viewed using a 4× EPlan objective (40×) to measure morphometric parameters of intestinal architecture.
For this purpose, three favorably orientated sections cut perpendicularly from villus enterocytes to the muscularis mucosa were selected from each animal and measurements were carried as follows: villous height (VH) was estimated by measuring the vertical distance from the villous tip to villous-crypt junction level for 10 villi per section; crypt depth (CD) (the vertical distance from the villous-crypt junction to the lower limit of the crypt) was estimated for 10 corresponding crypts per section.
Further parameters that were evaluated in this trial were measurements of pH and buffering capacity of the feeds, pH in the digestive tract of birds, intestinal microbiota and antioxidant status.
Statistical analysis
For the first trial and to investigate the effect of benzoic acid on turkeys’ performance, data was statistically analysed by analysis of variance using the PROC MIXED procedure of SAS (1989) with replication in time considered as a random effect. Linear and quadratic orthogonal contrasts were tested using the Contrast statement of SAS. Differences between treatments were declared significant at p<0.05.
For the second and third trials experimental data were analysed (SPSS v.17.00; SPSS, Inc., Chicago, IL, USA) as a randomized block design. Performance data were analysed by a one way ANOVA with initial body weight used as a covariate and the pen being the experimental unit. For data on antioxidant activity, intestinal morphology and bacteria loads individual birds were considered to be nested within pens and data were analysed by a nested ANOVA; in addition, data on antioxidant activity were analysed by a two way nested ANOVA with time and treatment being the experimental factors. As bacterial numbers were not normally distributed, they were log transformed to create a normal distribution prior to analysis. Bacteria load means are presented on transformed basis. Levene’s test was performed to check homogeneity of variances and Tukey’s test was carried out to assess any significant differences at a probability level of 0.05 among the experimental treatments.
RESULTS
In this study a basal diet (Table 1) was supplemented with various feed additives (one of the following compounds: benzoic acid, thymol, a mixture of essential oil blends (MEO) and a combination of benzoic acid and MEO (BMEO group) in order to study their effect on growth performance and certain health related parameters of turkey poults. Thus, three trials were conducted in order to identify the optimal level of benzoic acid (trial 1), the effect of thymol and MEO (trial 2) and the combined effect of benzoic acid and MEO (trial 3) on growth performance of turkey poults. In all trials, control groups were fed with the same basal diet (Table 1).
Effect of benzoic acid on growth performance
In the first experiment, benzoic acid was added in the basal feed at the levels of 300 mg/kg and 1,000 mg/kg resulting in the B300 and B1,000 groups respectively. The results of this trial are illustrated in Table 2 and as it shown the performance of turkey poults was improved (p<0.05) by the low dietary inclusion of benzoic acid. The pH values of both feed and caecal content decreased (p<0.05) following benzoic acid supplementation while no difference was noticed in the pH of the other parts of the digestive tracts. In the caecum, lactic acid bacteria populations were increased (p<0.05), and coliform bacteria decreased (p<0.05), following benzoic acid supplementation.

Effect of Benzoic acid (B) on performance of turkey poults, pH values in the digestive tract and intestinal microbiota in trial 1
Effect of thymol and MEO on growth performance
In the second trial, thymol or a mixture of essential oils (MEO) in the level of 30 mg/kg was added in the basal diet resulting in T30 and MEO30 groups respectively. As illustrated in Table 3, thymol supplementation did not change body weight gain or FCR compared to control feed. However, the mixture of essential oil (MEO) compounds improved (p<0.05), growth performance of turkeys compared to both the Control and thymol groups. Antioxidant status of turkeys was improved by both thymol and MEO supplementation (p<0.05). Dietary inclusion of the mixture of essential oil compounds increased (p<0.05) lactic acid bacteria populations and decreased (p<0.05) coliform bacteria population in the caecum. However, LAB or coliform counts were not affected in the crop or the ileum by dietary supplementation.
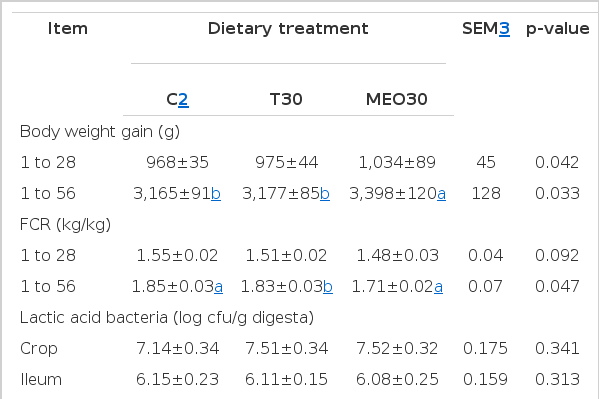
Effect of combination of benzoic acid and MEO on growth performance
In trial 3, the combine effect of benzoic acid and MEO on growth performance of turkey poults was examined. Therefore, CRINA poultry plus (which contains both benzoic acid and a mixture of essential oil compounds) was added in the basal diet resulting in BMEO group.
The results of this study are presented in Table 4 and as it is shown the addition of BMEO reduced the buffering capacity compared to control feed (p<0.05), improved growth performance, decreased the pH values of the caecal content, increased (p<0.05) lactic acid bacteria populations and decreased (p<0.05) coliform bacteria, in the caecum. BMEO inclusion elevated (p≤0.05) GSH-Px and GST activity and reduced (p≤0.05) MDA production in tissues compared to controls.
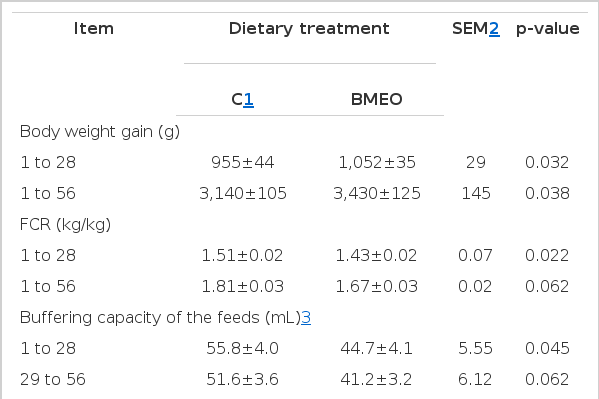
Performance results of turkey poults, intestinal microbiota, buffering capacity of the testing feeds, intestinal morphology and antioxidant status in trial 3
Therefore the mixture of benzoic acid and MEO (BMEO) resulted in higher growth performance compared to control; retained positive effects of both B300 and MEO30 groups and affected several gut related health parameters.
DISCUSSION
Although it has been common practice in animal farming to add organic acids to feeds for both their preservative and possible growth promoting effects (Falkowski and Aherne, 1984), literature data on the response of turkey poults to dietary benzoic acid are limited. In another study (Józefiak et al., 2010), very low (0.1%) dietary inclusion of benzoic acid resulted in increased BWG of chickens only in the first 2 weeks of fattening, while 0.2% supplementation decreased BWG in the grower (15 to 35 days) and the entire period of broiler chicken fattening. In a similar earlier study, a growth depressing effect of the benzoic acid at supplementation level of 0.25% to 0.75% was reported (Józefiak et al., 2006). In piglets higher (≥1%) supplementation levels of benzoic acid significantly improved performance compared to the controls in feed intake, BWG and FCR by 9%, 15%, and 6% respectively (Kluge et al., 2006). In contrast, in barrows (26 to 106 kg BW), addition of 1% benzoic acid to the diets did not improve weight gain and FCR, although it improved N-digestibility in the grower period (Buhler et al., 2006).
In similar trials, when chickens received 1% propionic acid in their diet, feed efficiency was improved 6% to 8% over the controls (Giesting and Easter, 1985). However, this improved feed efficiency was lost when the incorporation level of propionic acid increased to 2% and above. In another study (Watkins and Miller, 1983), addition of 0.5% and 1% calcium formate in broilers diets improved the live weight gain by 0.9% and 1.9%, respectively, compared to controls. On the other hand, administration of calcium formate with the feed in the range of 1.5% to 2.5% resulted in a dose dependent decrease of feed consumption and live weight gain. Feed conversion rate was improved in all calcium formate dosages up to 3.7%. Waldroup et al. (1995) reported that supplementation of broiler diets with a blend of formic and propionic acid at doses in the range 0.125% to 1% did not alter feed conversion rate or mortality and gave inconsistent results on body weight and feed consumption.
Part of the literature inconsistency might be probably attributed to the type and dose level of the organic acids applied and the health status of the animals. The response of broilers to in-feed antibiotics has been more marked when the performance level has been very low, the same may hold true for organic acids (Cave, 1984).
Literature inconsistency might be also due to differences in the buffering capacity value of the used diets. The buffering capacity value indicating the amount of acid needed to lower the pH of a feed to a certain value is important because it affects the course of digestion. High buffering capacity values in feeds pose higher risks for young animals, which have limited capacity to secrete gastric acid. When using feeds with high buffering capacity, the gastric pH will remain high, impairing protein digestibility. Undigested protein will reach the lower digestive tract where excessive protein fermentation may occur, leading to formation of toxic biogenic amines (Sturkie, 1976). In addition, poultry feeds with high buffering capacity may result in proliferation of harmful bacteria in the digestive tract. Hence, the buffering capacities of the diets used in the present study were examined (Tables 2 and 4).
It must be noted that the antibacterial activity of benzoic acid may be an effect both of the cation and the organic acid anion. Organic acids such as the formic or propionic acid are able to pass across the bacterial cell wall in their non-dissociated form. It was initially thought to be the non-dissociated acid molecule that was the antimicrobial agent but it is now recognized that after positive diffusion into the cell, the acid dissociates according to the cytoplasmic pH into anions and protons both of which could exert an inhibitory effect (Florou-Paneri et al., 2001).
The reason why, in broiler poultry nutrition, benzoic acid reduces the growth rate when fed at higher than 0.1% inclusions is not clear. It could be hypothesized that one of the reasons is its metabolic pathway – conjugation with ornithine. It was reported that the domestic fowl excreted benzoic acid and other aromatic acids as well as nicotinic acid, conjugated with ornithine (Nesheim and Garlich, 1963). Therefore, feeding benzoic acid in high levels could result in an arginine deficiency because dietary arginine is the source of ornithine in the fowl.
In the present study, FCR was improved by benzoic acid supplementation in the starter and finisher period. Better feed conversion of broiler chickens in the early stage of growth might be associated with the bacteriostatic effect of benzoic acid and less competition for nutrients between host and native microbiota. It has been shown that benzoic acid has antimicrobial properties, mainly because of its inhibitory effect on several microbial enzymes, in particular, a-ketoglutaric acid dehydrogenase and succinic acid dehydrogenase (Bosund, 1962). Knarreborg et al. (2002) demonstrated, in the batch culture conditions, that the benzoic acid and, to a lesser extent, fumaric acid exerted strong lethal properties towards lactic acid bacteria. The above authors suggested pH-dependent effects on coliform bacteria and lactic acid bacteria in stomach content and on coliform bacteria in content from the small intestine. The pH values used in the batch culture system by (Knarreborg et al., 2002) were supposed to mimic physiology of the pig stomach and so, in general, were lower than those observed in the chicken. In “in vivo” trials on piglets, dietary benzoic acid reduced anaerobic and lactic acid bacteria in the stomach. The same compound was found to decrease gram negative bacteria in the duodenum (Kluge et al., 2006). In the present experiment, coliform bacteria were reduced also in the ileum, although lower bacterial counts in the gastrointestinal digesta were not reflected in a better performance of the birds when fed 0.2% benzoic acid. This study has not shown significant effects of benzoic acid on the pH in the crop, gizzard and small intestine. This may be because of the fact that benzoic acid is a weak acid with high constant of dissociation, which might have been present in the stomach and small intestine largely in a non-dissociated form. It is well known that non-dissociated organic acids have antibacterial effects. They can passively diffuse through the bacterial cell wall; dissociate themselves when the pH is above the pKa. Then, the dissociated organic acids cause the internal pH reduction. The protons will be pumped out of the bacteria by an ATPase pump, which expends energy but the anions will accumulate in the cell and further become toxic (Jensen, 2001). The growth promoting effect seen in other species was not achieved by higher than 0.1% supplementation of this weak acid.
Essential oil compounds also possess strong antibacterial activities (Burt, 2004) and may serve as potential alternatives to AGP (Brenes and Roura, 2010). It is known also that phenol compounds may have synergistic effects with antibiotics or other antibacterial compounds such as organic acids or other phenolic compounds (Langeveld et al., 2013). Synergism via pharmacokinetic or physicochemical effects of phenolic compounds can be found even for substances that do not possess any pharmacological effects themselves (Langeveld et al., 2013). The results of this study show antibacterial changes in microbial fermentation as well as in selected bacterial populations may suggest that in lower parts of the GIT (i.e. caeca) benzoic acid and MEOs supplementation increased lactic acid bacteria. On the other hand, in the main absorption site in birds, which is ileum, a strong reduction in potentially pathogenic coliforms was recorded.
It has been reported that lactic acid producing bacteria may improve gastrointestinal function, feed digestibility and animal performance (Rehman et al., 2006). It is suggested that the establishment of Lactobacillus spp. prevents the colonization of pathogenic bacteria by competitive exclusion (van der Wielen et al., 2002). Lactobacilli and bifidobacteria compete against potential pathogens for nutrients and binding sites, thereby reducing the intestinal population of pathogens (Rolfe, 2000). Furthermore, lactobacilli and bifidobacteria produce organic acids and other bactericidal substances (Jin et al., 1998) all of which can suppress the colonization of the intestine by pathogenic bacteria.
Another finding of our study was that mucosal architecture was influenced by the combination of benzoic acid with essential oil consumption in terms of villus height (Table 4). The structure of the intestinal mucosa can reveal some information on gut health. Stressors that are present in the digesta can lead relatively quickly to changes in the intestinal mucosa, due to the close proximity of the mucosal surface and the intestinal content. Changes in intestinal morphology, such as shorter villi and deeper crypts have been associated with the presence of toxins (Yason et al., 1987) or higher tissue turnover (Miles et al., 2006). In the present study, significant increase in duodenal, jejunal and ileal villus height was noted. The intestinal villous can be regarded as the capacity of the bird to absorb nutrients from the feed. Longer villi are typically associated with excellent gut health and high absorptive efficiency. Cook and Bird (1973) reported a shorter villus and a deeper crypt when the counts of pathogenic bacteria increased in the gastrointestinal tract.
Another objective of our study was to investigate whether the sustained consumption of EOs would affect the antioxidant status of turkey tissues. We found that lipid oxidative stability, as well as glutathione-based enzyme activity, were significantly improved, both by thymol or EOs compounds at low supplementation level. The literature is abundant with evidence of in vitro and in vivo antioxidant activity of EOs compounds due to their ability to scavenge free radicals by single-electron transfer in vitro (Brenes and Roura, 2010). Certain soluble, low molecular weight polyphenolic compounds can be absorbed by the intestine, reaching the plasma and target organs. Although their levels in the circulation are low, with a reduced net absorption and relatively fast excretion half-lives, consumption of polyphenolics has been accompanied by increased total antioxidant activity (Jiang et al., 2007). Numerous studies have shown a postprandial antioxidant capacity of phenolic compounds from various feed stuffs (Jiang et al., 2007), however, when biomarkers of the redox status are measured after polyphenolic substance consumption, the results obtained are often contradictory (Papageorgiou et al., 2003).
We also observed an increased activity of the two antioxidant-enzymes in the EOs supplemented groups compared to control birds. The depletion in overall glutathione activity, observed within 5 days after refrigerated storage, indicated an ongoing process of oxidative stress in the examined tissues. Elevated antioxidant enzyme activities could be due to active induction of glutathione synthetic enzymes could be due to a sparing effect of glutathione by decreasing the oxidative load on the cells since selenium uptake was not changed. Although the latter seems more plausible, as MDA formation was found to be reduced in EOs supplemented groups, additional studies are required to determine which mechanism is responsible. It is possible that the antioxidant substances of EOs (Giannenas et al., 2011) are being utilised by the cells, thus sparing the intracellular antioxidant systems such as GSH and GSH-Px.
The results of the present study suggest that the combination of benzoic acid with essential oil compounds exerted a positive effect on the performance of turkey poults and improved gut integrity and intestinal microbiota. In vitro experiments revealed that the addition of benzoic acid reduced the buffering capacity of the feed offering a significant aim to birds to digest ingested feed. Further research is needed to establish this suggestion through more extensive investigation on feed digestion and growth performance of challenged birds with bacteria or protozoa that cause severe intestinal diseases.