Effect of Dietary Phytase Transgenic Corn on Physiological Characteristics and the Fate of Recombinant Plant DNA in Laying Hens
Article information
Abstract
The study aimed to evaluate the potential effects of feeding with phytase transgenic corn (PTC) on organ weight, serum biochemical parameters and nutrient digestibility, and to determine the fate of the transgenic DNA in laying hens. A total of 144 50-week-old laying hens were grouped randomly into 2 treatments, with 8 replicates per treatment and 9 hens per replicate. Each treatment group of hens was fed with diets containing 62.4% non-transgenic conventional corn (CC) or PTC for 16 weeks. The phytase activity for CC was 37 FTU/kg of DM, whereas the phytase activity for PTC was 8,980 FTU/kg of DM. We observed that feeding PTC to laying hens had no adverse effect on organ weight or serum biochemical parameters (p>0.05). A fragment of a poultry-specific ovalbumin gene (ov) was amplified from all tissues of hens showing that the DNA preparations were amenable to PCR amplification. Neither the corn-specific invertase gene (ivr) nor the transgenic phyA2 gene was detected in the breast muscle, leg muscle, ovary, oviduct and eggs. The digestibility data revealed no significant differences between the hens that received the CC- and PTC-based diets in the digestibility of DM, energy, nitrogen and calcium (p>0.05). Phosphorus digestibility of hens fed the PTC-based diet was greater than that of hens fed the CC-based diet (58.03% vs 47.42%, p<0.01). Based on these results, it was concluded that the PTC had no deleterious effects on the organ weight or serum biochemical parameters of the laying hens. No recombinant phyA2 gene was detected in muscle tissues and reproductive organs of laying hens. The novel plant phytase was efficacious in improving the phosphorus digestibility of laying hens.
INTRODUCTION
Phosphorus (P) is a mineral essential to energy metabolism and egg production in laying hens (Frost and Roland, 1991). However, in plant feedstuffs approximately 60% to 80% of P exists in the form of phytate and is not directly available to monogastric animals without phytase supplementation (Simons et al., 1990; Maenz, 2001). Poor availability of P for monogastric animals and consequent P-deficiency are ones of major constraints to animal production globally (Perney et al., 1993).
It has been well demonstrated that an exogenous supplement of microbial phytase is effective in improving phytate P digestibility in P-deficient diets (Ravindran et al., 1995; Carlos and Edwards, 1998). However, high production cost is one of the obstacles to the application of microbial phytase in non-ruminant animals’ diets (Zhang et al., 2000). Reducing feed costs at commercial farms is a constant concern for the poultry industry. With the progress of biotechnology, an Aspergillus niger-derived phytase expressed in the endosperm of corn has recently been developed (Chen et al., 2008). Phytase transgenic corn (PTC) will allow animal feed producers to avoid purchasing phytase and corn separately, and eliminate the need for mixing these two ingredients together.
It is hoped that PTC could be a feedstuff as well as an alternative to microbial phytase that would allow laying hens to digest more phytate P than is possible with non-transgenic conventional corn (CC). Except for the high phytase activity, fat content and non-phytate P content in PTC, its nutritional value is similar to that of CC (Gao et al., 2012). Recently, a series of studies comparing the feed safety of PTC with CC for animals have been conducted by our research group (Li et al., 2013; Ma et al., 2013). No differences have been observed in the laying performance or egg quality between hens consuming diets formulated with PTC or CC (Ma et al., 2013). Furthermore, no phytase gene fragments have been detected in the blood, liver, or feces of laying hens fed the PTC-based diet (Ma et al., 2013). The present study was to assess any differences in blood parameters or organ weight along with the digestive functioning of laying hens when fed a PTC- or CC-based diet. In addition, we wanted to verify a potential transfer of recombinant DNA to muscle tissues and reproduction organs of laying hens after consumption of a PTC- based diet.
MATERIALS AND METHODS
Animals and housing
This study was approved by the Animal Care and Use Committee of China Agricultural University. A total of 144 healthy Hy-Line Brown laying hens (BW = 1.80±0.10 kg, 50 weeks of age) were randomly allocated to one of the two dietary treatments with eight replicates each. Each of the eight replicates used three 45 cm×45 cm×45 cm stainless steel suspended cages containing three hens each. After a two-week adaptation period the hens were fed either the CC- or PTC-based diet for 16 weeks. The hens were housed in a completely enclosed, mechanically ventilated building, where they were maintained on a 16-h light schedule and allowed ad libitum access to diets and water. Room temperature was maintained at 25±2°C. Hens were monitored continuously for any variation in behavior.
Corn and experimental diets
The two experimental diets (Table 1) contained corn (62.4%) either from the CC and PTC. The CC and PTC were grown, harvested, and stored under the same environmental conditions (Yanqing, Beijing, China). The nutrient and energy contents of the corn varieties had been previously reported (Gao et al., 2012). The phytase activity for PTC was 8,980 FTU/kg of DM. The two treatment diets were formulated so as to contain adequate concentrations of all nutrients required for laying hens according to the National Research Council (NRC, 1994). To prevent contamination during diet preparation, both the control diet and the transgenic diet were mixed as single batches, the control diet first.

Collection and processing of samples
At the end of experiment, two hens from each replicate with similar live weights were selected for blood collection. Five milliliters of blood was harvested and centrifuged at 3,000 rpm for 10 min at 4°C. Serum was stored at −20°C for subsequent blood parameter analysis. After weighing, these hens’ throat was cut to allow bleeding and was then hung upside down for 3 min. All the surgical instruments were disinfected with 75% ethanol solution between animal. Immediately following sacrifice, the breast muscle, leg muscle, ovary, and oviduct tissues were excised, trimmed of any superficial fat or blood clots, weighed and collected. The samples were collected in separate sterile tubes, placed on ice, and stored at −80°C until analysis.
The relative weights of organs were calculated according to the following formula (Gao et al., 2010).
The levels of aspartate aminotransferase (AST), alanine aminotransferase (ALT), cholesterol (CHOL), alkaline phosphatase (ALP), blood urea nitrogen (BUN), creatinine (CRE), glucose (GLU), γ-glutamyl transpeptidase (γ-GT), lactate dehydrogenase (LDH), triglycerides (TRIG), uric acid (UC), calcium (CALC), inorganic phosphorus (IPHS), total protein (TP), albumin (ALB) and globulin (GLOB) were determined with the commercial diagnostic kits (Jiancheng Bio Co., Nanjing, China) and the automatic biochemical analyzer (CE-CX5, Beckman Corp., Fullerton, CA, USA).
Detection of the transgenic phyA2 gene
Three pairs of targeting a corn-specific invertase gene (ivr) gene, a poultry-specific ovalbumin (ov) gene and a foreign gene (phyA2) were obtained from Invitrogen (Beijing, China). The DNA extraction, primer sequences and polymerase chain reaction (PCR) cycling conditions were conducted as previously outlined by our group (Ma et al., 2013). The PCR products were separated with a 1.5% agarose gel in 1×TBE (89 mM Tris, pH 8.4; 89 mM boric acid; 2 mM EDTA) and was visualized using ethidium bromide staining.
Nutrient digestibility assay
In week 15, 5.0 g/kg of titanium dioxide (Ti) was added into the two diets as an indigestible marker for the calculation of nutrient digestibility. After a one-week adaptation period, six laying hens with similar live weights from each replicate were killed by intracardial injection of sodium pentobarbitone. The digestive tracts were removed immediately and the digesta of the ileal were expressed by gentle flushing with distilled water into plastic tubes. The digesta contents were pooled within a replicate, immediately frozen at −20°C, and then freeze-dried. Dried ileal digesta samples were ground using a mortar and pestle and then passed through a 0.5-mm sieve and stored at −4°C pending chemical analysis.
The dry matter, N, Ca, and P content of diet and digesta samples were analyzed according to AOAC (2000) procedures. All diet samples were analyzed for gross energy with an automatic adiabatic oxygen bomb calorimeter (PARR 1281, PARR Instruments, Moline, IL). The Ti content in the samples was analyzed using the procedure of Myers et al. (2004). Phytase activity in corn was determined according to the method of Engelen et al. (2001), with one phytase activity unit (FTU) defined as the quantity of enzyme that releases 1 μmol of inorganic P per min from 1.5 mM of sodium phytate at pH 5.5 at 37°C.
The apparent ileal nutrient digestibility of the diets was calculated according to the following formula (Ravindran et al., 2006):
Where, Tidiet/Tidigesta is the ratio of Ti in the diet and ileal digesta and Nutrientdigesta/Nutrientdiet is the ratio of nutrient in the ileal digesta and diet.
Statistical analyses
The experiment used a completely randomized design, and each experimental unit was a replicate. All data were analyzed using the one-way ANOVA analysis (SAS Institute, Inc., 2001). Results were expressed as standard error of mean (SEM). Effects were considered significant when p<0.05.
RESULTS AND DISCUSSION
During the past few decades, biotechnology has been widely used to produce genetically modified (GM) crops with input traits including insect resistance, herbicide tolerance, and nutritional improvements. In 2012, 30% of the canola, 35% of the corn, 81% of the soybeans, and 81% of the cotton planted globally were GM crops (James, 2012). However, the increased usage of GM crops as feed for meat-, egg- and milk-producing animals has led to public concern (Walsh et al., 2012). The GM plants intended for consumer consumption may pose a perceived risk to health along with the transfer of recombinant DNA from GM plants to livestock and livestock-derived products that are consumed by humans (Ma et al., 2013). Our previous study demonstrated that, except for the high phytase and fat content and lower phytate content in PTC, the nutritional values of PTC and CC are similar and within the normal ranges reported for CC (Gao et al., 2012). The performance of hens fed diets containing PTC, as measured by egg production and egg quality, is similar to that of hens fed diets formulated with CC (Ma et al., 2013).
Serum biochemical parameters and organ indices
In the present study, we found that the serum biochemical parameters and organ indices were similar for laying hens consuming diets formulated either with PTC or CC (Tables 2 and 3). This finding is consistent with those of previous studies in which no biologically significant differences in organ indices (McNaughton et al., 2008) or serum biochemical parameters (Appenzeller et al., 2009) were observed between animals fed diets prepared with GM or non-GM grain. This suggests that a diet prepared from PTC has no unintended effects on serum biochemical parameters and organ weight as compared to a CC-based diet.
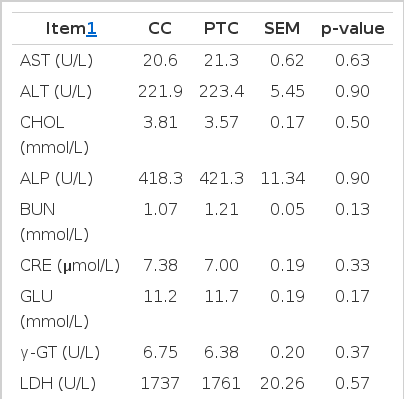
Serum chemistry values of laying hens fed diets containing conventional corn (CC) or phytase transgenic corn (PTC) for 16 weeks

Relative organ weights (g/kg BW) of laying hens fed diets containing conventional corn (CC) or phytase transgenic corn (PTC) for 16 weeks
Transgenic phyA2 gene detection in tissues of laying hens
The suitability of the PCR was verified through amplification of the corn-specific gene ivr and a poultry-specific fragment ov (Table 4). The ivr gene could be detected in all diet samples. In addition, results of PCR in all tissues were positive regarding the ov gene (Table 4). These results suggest that the PCR had been performed very well.
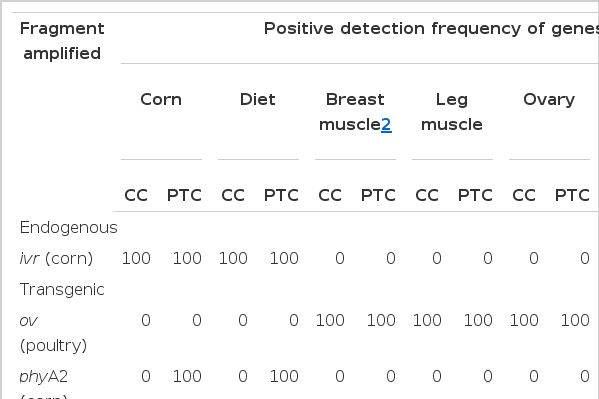
The PCR for the transgenic phyA2 fragment showed strong positive signals only for the PTC diet samples and no positive signals for the CC samples. No transgenic or corn-specific DNA was detected in any tissue samples such as breast muscle, leg muscle, ovary, oviduct, and eggs of the non-GM and GM groups. This may suggest that no corn-endogenous genes or transgenic fragments had been transferred to the hens’ tissues. These genetic materials were probably degraded too much to allow amplification in the muscle and reproductive organs of laying hens. In our previous study, detection of the recombinant phyA2 genes in the digesta of PTC-fed laying hens decreased rapidly as the digesta moved along the gastrointestinal tract of laying hens (Ma et al., 2013). No transgenic phyA2 gene was detected in the digesta of the ileal and rectum. In addition, we failed to detect any phyA2 transgenic fragments in the blood and eggs of laying hens fed diets containing PTC (Ma et al., 2013). These finding are consistent with previous studies investigating the fate of transgenic DNA in animals (Jennings et al., 2003; Phipps et al., 2003; Reuter and Aulrich, 2003; Aeschbacher et al., 2005). Walsh et al. (2011) stated that no transgenic or corn-specific DNA could be detected in the tissues of pigs (including heart, liver, spleen, kidney, and muscle) fed a GM corn-based diet.
Nutrient digestibility
The phyA2 phytase gene is expressed in the endosperm of PTC (Chen et al., 2008), unlike the microbial phytase which is exposed in the CC-based diets. Differences in expression systems might affect the releasing rate of the enzyme. The P-releasing efficacy of the novel PTC-derived phytase is another of our concerns.
Digestibility of nutrients in CC and PTC-based diets for laying hens are displayed in Table 5. Approximately 74% of the DM and 79% of the GE were utilized by hens (p>0.05). The apparent metabolizable energy content of the laying hens’ feed amounted to 13.05 and 13.24 MJ/kg of DM diet in the CC and PTC group, respectively. The N and Ca digestibility coefficients are not significantly influenced by corn variety (p>0.05). The P digestibility of the PTC group increased significantly by 22.40% as compared with that of the CC group (58.03% vs 47.42%; p<0.01).
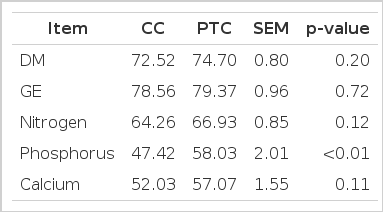
Effect of conventional corn (CC) or phytase transgenic corn (PTC) in the diet on the apparent ileal nutrients digestibility (%) of laying hens
The results of these analyses indicate that PTC had no adverse effects of the digestibility of DM, energy, N, and Ca of laying hens. These results are in agreement with findings published by Aulrich et al. (2001) and Aeschbacher et al. (2005). However, as expected, the effect of plant phytase supplementation on P digestibility was quite pronounced. The results show that a PTC diet can significantly improve P digestibility in laying hens. Similar observations were made by Nyannor and Adeola (2008), who demonstrated that using corn-based phytase (Escherichia coli-derived) in broiler diets was effective as microbial phytase in improving P digestibility. This clearly demonstrates that using PTC in diets can enhance P digestibility of poultry.
In conclusion, the feeding of laying hens with diets containing PTC does not affect organ weight or serum biochemical parameters as compared with hens fed a CC-based diet. No recombinant phyA2 gene was detected in muscle tissues and reproductive organs. Digestibility data from the study suggest that phytase expressed in PTC is efficacious in P-deficiency laying hens diets for the improvement of P utilization, which would minimize the need for supplemental P in diets.
ACKNOWLEDGEMENTS
This study was financed by the Genetically Modified Organisms Evaluation Major Projects (No. 2009ZX08011-005B-01), Scientific and Technical Supporting Programs (2011BAD26B03, 2013BAD10B01), Yangtze River Scholar and Innovation Research Team Development Program (NO. IRT0945), China Agriculture Research System (CARS-41-K15), and China Agriculture Research System-Beijing Team for Poultry Industry.