Polymorphisms in the Promoter Region of the Chinese Bovine PPARGC1A Gene
Article information
Abstract
The peroxisome proliferator-activated receptor gamma coactivator-1 alpha protein, encoded by the PPARGC1A gene, plays an important role in energy homeostasis. The genetic variations within the PPARGC1A gene promoter region were scanned in 808 Chinese native bovines belonging to three cattle breeds and yaks. A total of 6 SNPs and one 4 bp insertion variation in the promoter region of the bovine PPARGC1A gene were identified: SNP -259 T>A, -301_-298insCTTT, -915 A>G, -1175 T>G, -1590 C>T, -1665 C>T and -1690 G>A, which are in the binding sites of some important transcription factors: sex-determining region Y (SRY), myeloid-specific zinc finger-1 (MZF-1) and octamer factor 1(Oct-1). It is expected that these polymorphisms may regulate PPARGC1A gene transcription and might have consequences at a regulatory level.
INTRODUCTION
The peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PPARGC1A, also known as PGC-1α) protein, encoded by the PPARGC1A gene, plays an important role in energy homeostasis. PGC-1α is a metabolic switch, which convergently regulate metabolic pathways through its pleiotropic interactions with nuclear receptors (NRs) and other non-NR transcription factors. PGC-1α transcriptionally activates a complex pathway of mitochondrial biogenesis, lipid and glucose metabolism (Lin et al., 2005; Puigserver, 2005). The expression of PGC-1α is enriched in tissues where mitochondria are abundant and oxidative metabolism is active, such as brown adipose tissue (BAT), the cardiac and skeletal muscles (Puigserver et al., 1998).
Independent studies in humans have identified a common hPPARGC1A polymorphism, Gly482Ser, associated with type 2 diabetes (Barroso et al., 2006; Bhat et al., 2008; Povel et al., 2010; Geloneze et al., 2012). The most common polymorphism c.1892-19T>C in bovine PPARGC1A intron 9 was studied for the associations with milk production and milk-fat composition traits (Weikard et al., 2005; Khatib et al., 2007; Komisarek and Dorynek, 2009; Schennink et al., 2009; Kowalewska-Luczak et al., 2011). Several other polymorphisms in the bovine PPARGC1A gene have been identified and studied for the association with meat quality and carcass phenotypes (Soria et al., 2009; Ryu et al., 2012).
As an important component of the world’s bovine population, the Chinese native population possesses very abundant genetic resources. However, up to now, there was no information about the genetic variation of PPARGC1A gene promoter in Chinese indigenous bovine breeds. Therefore, a scan of the genetic variations within the PPARGC1A gene promoter in Chinese native bovines, should provide important information on bovine genetic resources that could be useful for selection and breeding through marker assisted selection (MAS).
MATERIAL AND METHODS
Bovine populations, genomic DNA isolation and pooling
Genomic DNA samples were obtained from 808 bovines belonging to three cattle breeds and yaks. Three Chinese indigenous cattle breeds, Jiaxian (JX, n = 148), Nanyang (NY, n = 278), Qinchuan (QC, n = 325) were investigated. The Jiaxian cattle were from the breeding farm of Jiaxian cattle (Jiaxian county, Henan Province, China); the Nanyang cattle were from the breeding farm of Nanyang cattle (Nanyang city, Henan Province, China); the Qinchuan cattle were from the reserved farm (Fufeng county, Shaanxi Province, China), the breeding and seed centre of Qinchuan cattle (Yangling District, Shaanxi Province, China). The yaks (n = 57) were from the breeding farm of yaks (Datong county, Qinghai province, China). DNA was isolated from 1 ml whole blood samples according to Sambrook and Russell (2001). DNA of 50 to 100 randomly chosen individuals from the same breed were mixed to one DNA pool (Bansal et al., 2002).
Primers designing
Six primer pairs were designed with the Primer V5.0 software based on NCBI database (GenBank accession number AC_000163) to amplify the promoter region of the bovine PPARGC1A gene. Detailed information about oligonucleotide primers, amplicon sizes and corresponding annealing temperatures is given in Table 1.

Primer information of bovine PPARGC1A gene
Single nucleotide polymorphisms (SNPs) detection
The genomic DNA pool samples were used as templates. Each amplification reaction was carried out in a total volume of 25 μl in the presence of 50 ng bovine genomic DNA pool as template, 0.5 μM of each primer, 1× buffer (including 1.5 mM MgCl2), 200 μM dNTPs (dATP, dTTP, dCTP and dGTP), and 0.625 U Taq DNA polymerase (MBI, Vilnius, Lithuania). The cycling protocol was 4 min at 95°C, 35 cycles of denaturing at 94°C for 30 s, annealing at the optimized temperature for 30 s, extending at 72°C for 30 s, with a final extension at 72°C for 10 min. All PCR products were then sequenced using ABI 3730 sequencer (ABI, Foster City, CA, USA), in both the forward and reverse directions, and the sequences were imported into the Megalign software module of DNASTAR (version 4.0) and BioXM software (version 2.6) to search for SNPs.
SNP genotyping
The genomic DNA samples were used as templates to individually and specifically amplify the promoter region of PPARGC1A. SSCP analysis of PCR fragments obtained above was performed on electrophoresis units (Bio-Rad, Amersham), coupled with a refrigerated system. Aliquots of 5 μl PCR products were mixed with 5 μl denaturing solution (95% formamide, 25 mM EDTA, 0.025% xylene-cyanole and 0.025% bromophenol blue), heated for 10 min at 98°C and chilled on ice. Denatured DNA was subjected to PAGE (80×73×0.75 mm) in 1×TBE buffer and constant voltage (200 V) for 2.5 to 3.0 h. After electrophoresis, gels were stained with 0.1% silver nitrate and visualized with 2% NaOH solution (supplied with 0.1% formaldehyde) according to Zhang et al. (2007). After the polymorphisms were detected, three PCR products of each different electrophoresis pattern were sequenced in both directions in an ABI 3730 sequencer (ABI, Foster City, CA, USA) and analyzed with BioXM software (version 2.6).
Data analyzing
Allele frequencies were determined for each breed by direct counting. The program MatInspector (http://www.genomatrix.de) was performed to identify any known transcription factors that may potentially bind to polymorphic promoter sequences in the TRANSFAC database.
RESULTS AND DISCUSSION
In the present study, genomic DNA of all bovine breeds was successfully amplified using primer pairs for the promoter region of the bovine PPARGC1A gene. A total of 6 SNPs and one 4 bp insertion variation in the promoter region of bovine PPARGC1A gene were identified. In the PCR product primer1, the SNP -259 T>A and -301_-298insCTTT were found; in primer3, -915 A>G; in primer4, -1175 T>G; in primer5, -1590 C>T, -1665 C>T and -1690 G>A. All these polymorphisms are numbered relative to the adenine of the translation start codon. The sequencing results and SNPs found are shown in Figure 1. The nature and the distribution of PPARGC1A promoter variations are shown in Table 2.

Sequencing results and polymorphic sites found in the promoter region of the bovine PPARGC1A gene in Chinese breeds. Adenine of the start codon ATG is counted as +1 (GenBank accession No. AC_000163).
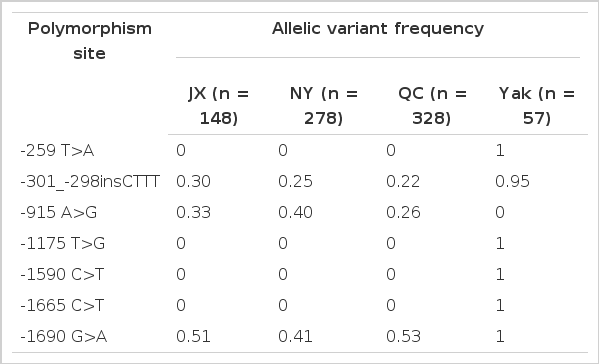
The SNPs identified in the promoter region of the bovine PPARGC1A gene in Chinese breeds and allele frequencies of the SNPs
The SNPs -259 T>A, -1175 T>G, -1590 C>T and -1665 C>T occurred only in the Yak population. Conversely, the SNP -915 A>G occurred only in cattle breeds (JX, NY and QC), while -301_-298insCTTT and SNP -1690 G>A occurred both in Yak and cattle populations. The genetic differences of PPARGC1A promoter in the present study give support to the notion that the yak is an independent genus, Poephagus. This notion was also supported by the reports from Zhang (Zhang et al., 2010; Zhang et al., 2011). The discrepancy in different populations was probably caused by the following two possible reasons. First, the yak was an ancient animal species with many wild bovine characteristics, which did not undergo much artificial selective breeding. Meanwhile, the Jiaxian, Nanyang and Qinchuan cattle, three of the most representative indigenous bovine (Bos taurus) breeds in China, are draft and beef dual-purpose cattle breeds which have been selected for beef production in the past decades. Second, yaks must survive in harsh environments (low temperature, hypoxia with high altitude). So, some variations may only exist in yaks as a result of adaptation their environment.
Polymorphisms in promoter region may affect gene transcription through changing the affinity between the transcription factors and promoter sequences. To investigate the potential effect that some of the polymorphisms could have on gene transcription, the program MatInspector (http://www.genomatrix.de) was performed in the TRANSFAC database to identify any known transcription factors that may potentially bind to polymorphic promoter sequences. The -301_-298insCTTT makes the recognition sequence of sex-determining region Y (SRY) repeated. The SRY locus on the mammalian Y chromosome is the developmental switch responsible for testis determination. Inconsistent with this important function, SRY has been demonstrated to modulate the catecholamine pathway, so it should have functional consequences in the central and peripheral nervous system (Turner et al., 2011). The -915 A>G makes the recognition sequence of myeloid-specific zinc finger-1 (MZF-1) disappeared. The zinc finger gene MZF-1 is preferentially expressed in primitive hematopoietic cells and plays an important role in regulating myelopoiesis (Hromas et al., 1995). In this polymorphic site, the yak population was in homozygous AA status, which preserves the recognition sequence of MZF-1. These traits are crucial for yaks to survive in severe environments. The -1665 C>T is in the core recognition sequence of octamer factor 1 (Oct-1). It is now known that Oct-1 controls not only housekeeping but also numerous tissue-specific genes. The latter include the genes for interleukins (IL) 2, 8, 3, 5; the granulocyte-macrophagal colony-stimulating factor, immunoglobulins α; the light and heavy chains of immunoglobulins; the endocrine-associated Pit-1 gene; the genes for gonadoliberin, prolactin, the thyroid transcription factor 1, and thyrotropin (Sytina and Pankratova, 2003). It is expected that these polymorphisms regulate PPARGC1A gene transcription and might have consequences at a regulatory level, but there are no data available to suggest the interaction between the variations and the PPARGC1A promoter activity until now. The potential biological function of the mutation in bovine PPARGC1A promoter remains to be further studied.
Acknowledgements
This study was supported by the National Natural Science Foundation of China (No. 30972080 30901023), Program of National Beef Cattle Industrial Technology System (No. CARS-38), Agricultural Science and Technology Innovation Projects of Shaanxi Province (No.2012NKC01–13).