Effects of Monensin on Metabolism and Production in Dairy Saanen Goats in Periparturient Period
Article information
Abstract
This trial evaluated the effects of dietary supplementation with monensin sodium on dry matter intake, metabolic parameters and milk yield and milk composition of dairy Saanen goats in the periparturient period. Twelve Saanen pregnant dairy goats were assigned to a treatment and a control group. Saanen goats were fed monensin as its 10% sodium salt in the amount of 33 mg/kg of total dry matter intake during 30 d before parturition till d 42 in milk. Blood samples were collected from each goat at d 30, 15 and 7 before expected kidding time and also in d 1, 7, 13, 19, 21, 28, 35 and 42 postpartum. The serum concentrations of β-Hydroxybutyrate (BHBA), non-esterifed fatty acid (NEFA), calcium, magnesium, inorganic phosphorus, cholesterol, triglyceride, urea, total protein, albumin and glucose and aspartate aminotransferase (AST) activity were determined. The BHBA concentration significantly decreased in goats, which received monensin in comparison to controls in the postpartum period (p = 0.049). The concentration of sodium (Na) was significantly influenced by monensin treatment in the postpartum period (p = 0.048). In addition, the difference in dry matter intake was highly significant between the two groups during the pre-partum period. Controls had more dry matter intake (DMI) than treatment goats (p = 0.0001). Mean 3.5% fat corrected milk production was not influenced by monensin treatment. However, milk fat percentage was significantly decreased in monensin treated goats (p = 0.0017).
INTRODUCTION
Ketosis, as seen in dairy cows (ie, acetonaemia) or sheep (ie, pregnancy toxaemia), also may be encountered in goats (Harwood, 2004). Ketosis/pregnancy toxemia occurs in a doe (before or after kidding) because of an inability to consume enough feed to meet her needs. After kidding ketosis results from the doe producing higher milk yields than her body can sustain. The importance of ketosis in Saanen goats, which produce more milk, is greater than the other breeds of goats.
One potential way to help alleviate negative energy balance may be through the inclusion of ionophores. Ionophores act to alter the rumen microflora through ion transfer across cell membranes. One of the most commonly used ionophores is monensin (Tyler et al., 1992). Monensin has approved for use in lactating cows in many countries (Duffield et al., 2008). Monensin is a carboxylic polyether ionophore that is produced by the fermentation of Streptomyces cinnamonensis. It changes the rumen microflora, resulting in a decrease of the microbes that produce acetate and butyrate and increasing the microbes that produce propionate, the gluconeogenic precursor (Bergen and Bates, 1984). Monensin treatment in lactating dairy cattle reduced the blood concentrations of β-Hydroxybutyrate (BHBA), acetoacetate, and non-esterifed fatty acid (NEFA). These findings follow a logical pattern of improved, coordinated energy metabolism, which is particularly beneficial for the transition dairy cow in providing a more positive adaptive response to the challenges of lactation (Duffield et al., 2008). Furthermore, inclusion of monensin in lactating cow diets resulted in both increased milk yield and decreased DMI, and improved milk production efficiency (Duffield et al., 2008). Supplementation of monensin to pregnant ewes during prepartum period had a markedly positive influence on energy indicators in late pregnancy (Taghipoor et al., 2011). No milk withholding time and no pre-slaughter withdrawal period is required for monensin (Dick et al., 1996; Bagg et al., 2005).
The 1st objective of the study herein was to evaluate the metabolic effect of monensin administered 30 d before parturition till 42 d after parturition on serum biochemistry parameters in the weeks pre- and post-parturition in dairy Saanen goats. The 2nd objective was to investigate the effects of monensin on dry matter intake, milk production and milk composition of Saanen goats.
MATERIALS AND METHODS
Study design, animals and diets
Twelve Saanen does were enrolled in an experimental trial. The does were synchronized for estrus by using two injections of PGF2α 11 d apart. Rams were introduced to does 3 d after the second dose. The rams remained with does for one week. Pregnancy was diagnosed by ultrasonography on d 60 and 80 after mating using a portable B-mode ultrasound scanner. The number of fetuses was recorded during the ultrasonographic diagnosis of pregnancy. The health of all goats was examined by clinical examination and CBC analysis before the beginning of the study. The does were treated by Ivermectin and Triclabendazole for parasite control. Thirty days before expected time of parturition, the does were blocked by age and assumed fetal number and randomly divided into two groups. Animals in treatment group (N = 6) received monensin as its 10% sodium salt at a rate of 33 mg/kg DM that was mixed in their daily diet from 30 d before expected kidding time to d 42 postpartum. Goats in control group (N = 6) did not receive monensin in their diet. All animals transferred to individual pens and were fed twice daily and fresh water was available ad-libitum. Both groups were fed the same pre-partum and post-partum rations. The diets were formulated to meet all nutrient requirements for pregnant does at pre-partum and lactating does during the postpartum period (NRC, 2007). The ingredients of prepartum and postpartum diets are shown in Table 1. Daily diet of each goat was weighed prior to feeding and feed refusals were measured by digital scale weigher for accurate estimation of daily DMI of each animal.

Ingredients and chemical composition of the experimental diets
The does were weighed on d 30 and 15 prior to parturition and on d 1, 15 and 30 postpartum. The kids were weighed at birth time.
Sampling and biochemical analysis
Blood samples were collected from the jugular vein of each goat at d 30, 15 and 7 before expected kidding time and also in hour 12 and d 7, 13, 19, 21, 28, 35 and 42 postpartum. Because of the difficulty in predicting parturition day, samples taken prior to kidding were grouped as d 29 prior to parturition (SD = 3.72), d 14 (SD = 2.17), and d 6 (SD = 1.97). All blood samplings were taken 3 h after first daily meal at the same time of day for all animals. The blood specimens were collected into 10 ml vacuum tubes and were immediately transferred to laboratory with an unbroken cold chain. Each sample was centrifuged and its serum was harvested, frozen and stored at −20°C until the time of analysis. The concentrations of BHBA and NEFA were determined by a D-3-hydroxybutyrate kit and a NEFA Kit (Randox Laboratories Ltd, Ardmore, UK). Calcium, magnesium, inorganic phosphorus, cholesterol, triglyceride, urea, total protein, albumin and glucose concentrations and AST activity were determined by an automated biochemical analyzer (Biotecnica instruments, BT 1500, Rome, Italy) using commercial kits (Parsazmoon, Tehran, Iran). Sodium (Na) and potassium (K) concentrations were determined by flame photometery (Elico, Model CL 361, India). Blood chemistry analysis was based on the following colorimetric methodologies, Total protein: Biuret reaction (Tomas, 1998), Albumin: Bromocresol green (Johnson et al., 1999), UREA: urease UV (Tomas, 1998), Cholesterol: CHOD-PAP (Deeg and Ziegenhorn, 1983), Magnesium: xylidyl blue-EGTA (Ratge et al., 1986), Phosphorus: phosphomolybdate UV (Daly and Ertingshausen, 1972), Calcium: arsenazo (Baginski et al., 1973), Glucose: glucose oxidase (Barham and Trinder, 1972), Triglyceride: GODPAP (Cole et al., 1997), AST: IFCC (Bergmeyer et al., 1986). BHBA (McMurray et al., 1984) and NEFA (DeVries et al., 1976) were measured with commercial colorimetric Kits. Measurement of data accuracy was done with the use of Control serum (Randox control sera, Antrim, UK). All parameters showed a within-run impression of less than 10% (Total protein: 0.95%, Albumin: 0.91%, UREA: 5.41%, Cholesterol: <3%, Magnesium: 1.2%, Phosphorus: 1.87%, Calcium: 1.45%, Glucose: <3%, Triglyceride: <3%, AST: 2.5%, BHBA: <5% and NEFA: <5%).
CBC analysis was performed in d 30, 15 and 7 before parturition and d 3, 10, 20 and 35 postpartum for monitoring health status of animals. Blood specimens for CBC analysis were collected in EDTA containing tubes and analyzed by an automated cell counter (Nihon Kohden, Tokyo, Japan).
After parturition, all animals were machine-milked twice daily and the milk yield of each doe was recorded. Also in d 5, 20, 35 and 50 of lactation, milk specimens were collected and frozen for analysis of milk composition. Milk composition analysis was done by milk analyzer (Lactostar, TMS Ltd., Germany).
Data management and statistical analysis
The data were checked for errors and compared with written reports; outliers were rechecked to ensure that values were accurate. Because serum metabolites were measured over time, a repeated measures approach using ANOVA with Mixed linear models (PROC MIXED) was used in SAS (The SAS system for Windows, Version 8.2, Cary, NC, USA). All outcome variables were screened for normality by visual assessment of the distributions and calculation of kurtosis and skewness. The distributions of cholesterol, albumin, total protein, calcium, inorganic phosphorus, magnesium, sodium and potassium were normal. Serum concentrations of BHBA, NEFA, glucose, triglyceride, urea, and AST were skewed to the right. Therefore, a logarithmic transformation was performed on each of these dependent variables, which resulted in a normal distribution. Goat variables included treatment, age, and the number of fetuses, and body weight at sampling time (except for the models in which body weight was the outcome). Age as a covariate variable was classified as 1, less than 3 yrs old and 2, equal to or more than 3 yrs. Goats were classified as thin, a body weight of greater than 45 kg as fair.
The metabolites and DMI were separately analysed at pre-partum and postpartum periods. Body weight, milk production, milk fat percentage, milk fat yield, milk protein percentage, and milk protein yield were considered as outcome variables. Outcome variables were treated as repeated measures within goat in PROC MIXED. Age as a covariate variable was considered for milk yield and milk components. Several covariance structures were evaluated for each outcome: compound symmetry, unstructured, auto regressive order 1, autoregressive heterogeneous order 1, topelitz and topelitz heterogeneous. The covariance structure that resulted in Akaike’s information criterion (AIC) closest to zero was used (Wang and Goonewardene, 2004).
Preliminary Mixed models were used to screen for significant variables and their interactions with monensin. The interaction term was removed from the model when p> 0.25. A final model for each outcome variable was built using manual backward elimination procedure with a p-value of <0.05 for a partial F-test. All reported values are least squares means.
RESULTS
Animals, twinning, body weight of kids, and feed intake
The average age of goats was 3.92±3.01 yr (range: 2 to 10 yr) and 3.58±3.22 yr (range: 1.5 to 10 yr) in monensinfed and control groups, respectively. The mean body weight of goats 30 d ante-partum was 52.45±11.24 kg in monensin treated group and 41.57±9.73 kg in controls. One day postpartum, does in treatment group weighed 51.05±11.74 kg and does in control group weighed 41.60±10.70. At 30 d after kidding the mean body weight of treatment and control groups were 46.70±10.69 kg and 37.97±6.39 kg, respectively. Although the body weight difference between two groups was remarkable, it was not statistically significant. In addition, differences of age and weight gains during samplings between groups were not statistically significant.
Four out of 6 does in treatment group and two out of 6 controls bore twin kids. The mean body weight of kids in treatment group was 3.00±0.214 kg and in control group was 2.830±0.222 kg. The difference of body weight of neonatal kids between groups was not significant.
Treatment group fed 1,333.66±39 g dry matter/d and control group fed 1,481±30 g dry matter/d during 30 d pre-partum. The difference of dry matter intake was highly significant (p = 0.0001) between two groups pre-partum (Table 2). The controls with less body weight received more DMI. Therefore, body weight was not an influencing factor on the significant difference between two groups. However, the mean daily dry matter intake was 2,060.4±53 g in monensin group and 2,048.6±58 g in controls during 42 d in milk (p = 0.84).

Comparison of least square of means (LSMs) and standard errors (SE) of dry matter intake between Saanen does receiving monensin and the controls which did not received it
Serum biochemical analysis
No significant effect of monensin was seen on the concentration of BHBA pre-partum (Table 3). However, the concentration of BHBA was significantly affected by monensin treatment in the postpartum period (Table 4). The BHBA concentration significantly (p = 0.049) decreased in does which received monensin in comparison to the control group after parturition (Figure 1).
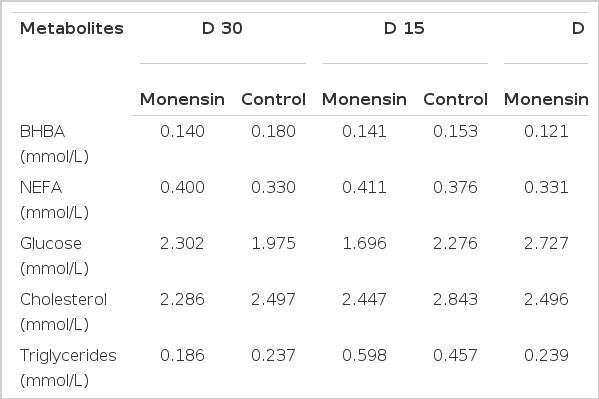

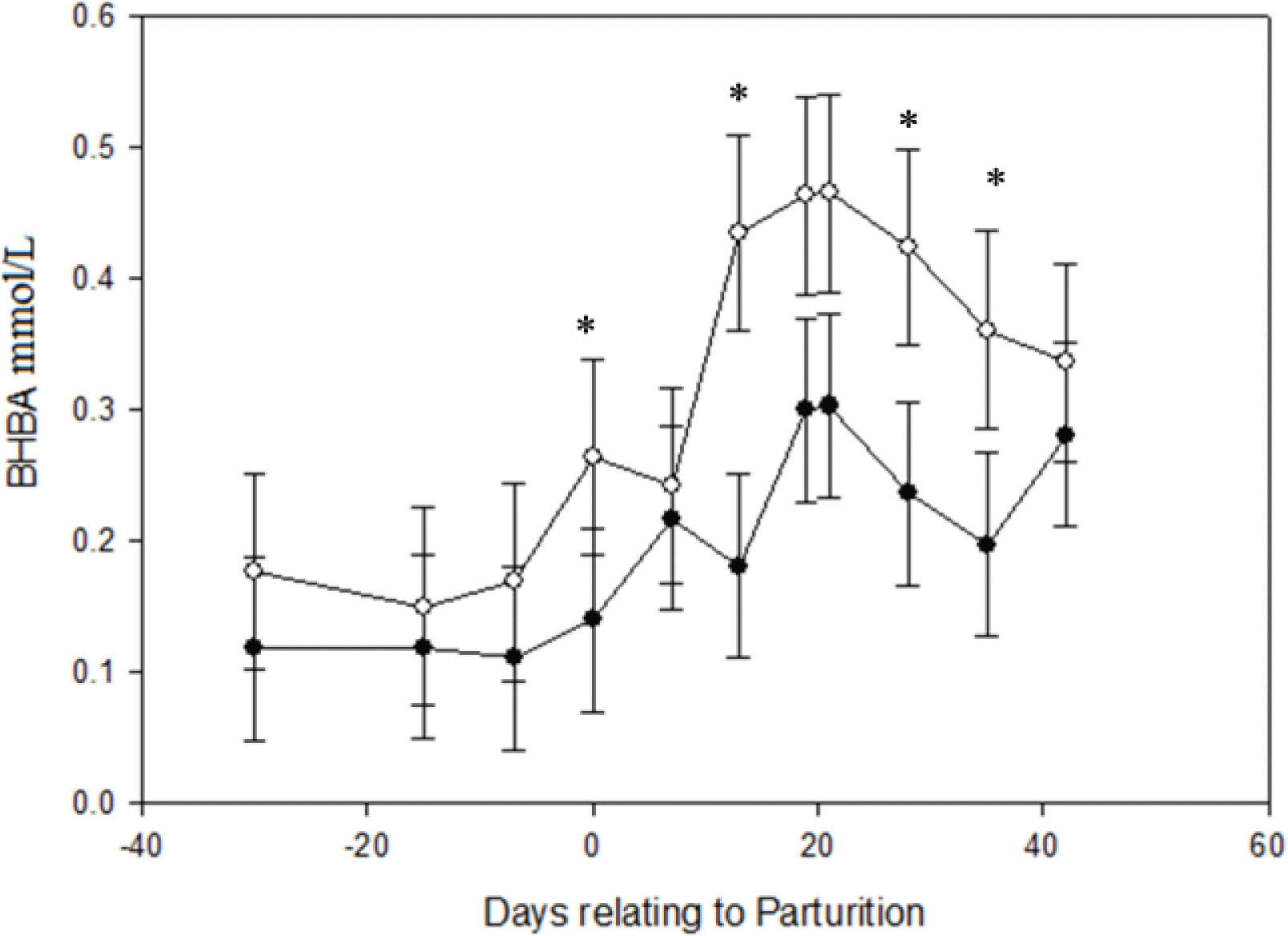
Least square of means values and standard errors for serum BHBA (mmol/L) concentration at 30 d before parturition till 42 d in milk in Saanen goats receiving monensin (●), and not receiving monensin (○). Significant time×treatment interactions (p<0.05) on d 1, 13, 28 and 35 were showed by an asterisk.
No significant effect of monensin was seen on the concentrations of NEFA, glucose, cholesterol, triglyceride and activities of AST at prepartum and postpartum periods.
The concentration of Na was significantly influenced by monensin treatment (p = 0.048) after parturition (Table 4). No significant monensin effects were found for albumin, total protein, urea, calcium, inorganic phosphorus, magnesium and K in pre- and post-partum periods (Tables 3 and 4).
Milk production and components
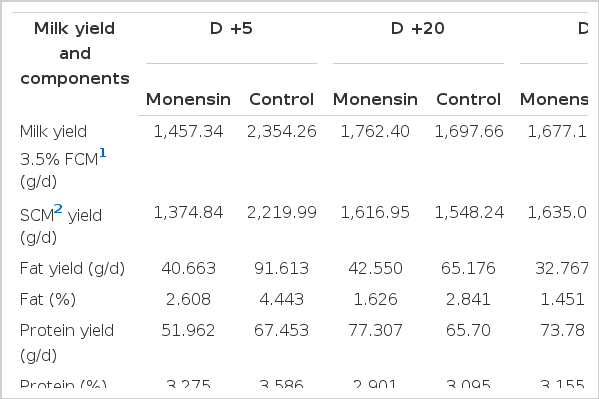
Mean 3.5% fat corrected milk production was not influenced by monensin treatment. Milk fat percentage was significantly (p = 0.0017) decreased in monensin treated goats in comparison with the control group (Figure 2). The milk fat and protein yields and percentages of milk protein, milk lactose and milk dry matter (g/daily) was not influenced by monensin (Table 5).

Least square of means values and standard errors for milk fat percent during 50 d in milk in Saanen goats receiving monensin (●), and not receiving monensin (○). Significant time×treatment interactions (p<0.05) were showed by an asterisk.
Comparison of least square of means (LSMs) and standard errors (SE) of milk composition parameters between Saanen does receiving monensin and controls not receiving it
DISCUSSION
The main objective of the study presented here was to explore potential effects of monensin supplementation during the last month of pregnancy and 42 d of lactation on serum biochemistry and milk production of dairy Saanen does. Comparison of serum biochemical constituents showed significant differences of BHBA and Na concentrations between treatment and control groups. To the knowledge of the authors, there is only one peer reviewed paper on the effect of monensin on milk composition and ruminal volatile fatty acids present in lactating goats (Brown and Hogue, 1985).
The administration of monensin to pregnant Saanen goats during 30 d before expected parturition till 42 d of lactation significantly reduced BHBA concentration when compared with a control group in post-partum period. This finding is consistent with results from monensin supplemented dairy cows (Petersson-Wolfe et al., 2007; Duffield et al., 2008). The beneficial effect of monensin on negative energy balance was seen in pre-partum period in ewes (Austin and Wilde, 1985; Taghipoor et al., 2011); however, the hypoketonemic impact of monensin was more prominent in the lactation period during d 1, 13, 28 and 35 postpartum in dairy Saanen goats. This finding is similar to the results obtaining in dairy cows (Duffield et al., 2008). It is reasonable to expect the effects of monensin on energy metabolism to occur in prepartum ewes in contrast to dairy goats where the greater effects were seen in the post-parturition period. Negative energy balance appears to be related to the glucose demands of lactation in dairy goats similar to that of dairy cows. However, in pregnant ewes the glucose utilization of fetal-placental unit is the major glucose demanding mechanism as eighty percent of fetal growth occurs during the last 6 wks of gestation in sheep (Edmondson and Pugh, 2009).
One of the major beneficial mechanisms of monensin on energy metabolism is increasing of propionate production in the rumen by inducing an alteration in rumen microflora (an increase in gram negative bacteria and a decrease in gram positive bacteria). While propionate is the major precursor of gluconeogenesis, more propionate production leads to a better function of this metabolic pathway. The end product of this pathway is glucose. Since the ketone bodies suppress the endogenous glucose production, monensin can indirectly cause elevation of blood glucose concentration by lowering ketone bodies (Schlumbohm et al., 2004). The increased concentration of glucose was not seen in monensin-fed goats in the present study. The finding that glucose concentration was increased with monensin has not been consistently reported in the literature. The lack of a significant effect of monensin on glucose concentration most likely is a function of the response being small and generally requiring a larger sample size to effectively assess the glucose response (Duffield et al., 2008).
Monensin could also reduce BHBA by reducing butyrate in the rumen, which is converted to BHBA in the rumen epithelium. A rise in BHBA occurs after meals, which is believed to be a result of an increase in conversion of butyrate into BHBA in the rumen epithelium (Plaizier et al., 2005).
Other energy and protein biochemical indicators particularly NEFA and cholesterol were frequently not significant possibly due to the large number of subjects needed to show statistically significant effect (Duffield et al., 2008).
There was a significantly higher Na concentration in goats receiving monensin. The basic mode of action of monensin and other ionophores is to modify the movement of ions across biological membranes. This basic function has yielded the name ionophore or “ion bearer” (Schelling, 1984). The effectiveness of monensin to cause Na entry into cells has been well established (Smith and Rosengurt, 1978), and other reported cellular ion changes may be secondary to the primary Na effect (Schanne et al., 1979). Scott et al. (1984) showed that monensin increased apparent absorption of sodium in steers. In agreement with the present study, Lasalocid (another ionophore) supplementation significantly increased serum Na concentration during late pregnancy and during lactation in Rambouillet ewes (Shetaewi and Ross, 1991). However, this effect of monensin on Na in dairy cows and ewes was not reported in previously published papers (Austin and Wilde, 1985; Taghipoor et al., 2011). The biological significance of the finding is not obvious and it might be a random effect. Furthermore, the serum Na concentration remained within the normal range throughout the experimental period.
In the current study DMI was lower for Saanen does receiving monensin than does serving as controls (p = 0.0001) during the prepartum period. However DMI was not significantly different between two groups at post-partum period. Although the body weight difference between two groups was remarkable, it was not statistically significant. It is noteworthy that the controls with less body weight received more DMI. Therefore, body weight was not an influencing factor on significant increase of DMI in controls versus treatment group. On the other hand, body weight was considered as a covariate in statistical analysis of serum metabolites and DMI.
The impact of monensin on DMI in dairy cattle is inconsistent in the literature. A meta-analysis of the effect of monensin on production outcomes in dairy cattle, conducted using 77 trials, indicated significantly decreased dry matter intake by 0.3 kg (Duffield et al., 2008).
The work by Sauer et al. (1989) reported a decrease in DMI in cows treated with 33 ppm of monensin during early lactation. Wagner et al. (1999) also showed decreased DMI during the whole lactation period with monensin at the level of 16 and 24 ppm. However, in one review, the authors found that 8 of 12 studies on ionophores reported no significant difference for DMI (Ipharraguerre and Clark, 2003).
Our results showed significant DMI decrease only at pre-partum and not at post-partum period. Schelling (1984) indicated that severe feed depression was seen when abruptly introducing cattle to monensin, which reduced feed intake by as much as 16%. Studies indicate depressions of less than 5% after 112 d on feed (Schelling, 1984).
The finding on DMI during the post-partum period is also consistent with results obtained by Brown et al. (1985) in French Alpine does and results of Petersson-wolfe et al. (2007) in cows. Petersson-Wolfe et al. (2007) have reported no significant impact of monensin delivered with controlled- release capsule and as feed additive on DMI in dairy cows in the transition period. In a study on the effect of monensin in lactating goats, no significant effect of monensin was seen on feed intake (Brown and Hogue, 1985).
The influence of monensin on digestibility is variable and the response is often unfavourable during adaptation periods. Several studies suggest that monensin may initially produce a negative effect followed by a slight positive effect. Simpson (1978) found that monensin decreased cellulose digestibility when no adaptation time was allowed, but reports indicate that monensin had no effect on cellulose digestibility when animals had been adapted to monensin for 21 d (Dinius et al., 1976). A reduction in dry matter and ADF digestibility was observed when lambs were fed monensin for only 10 d, but after an additional 29 d of adaptation, there was no significant difference (Poos et al., 1979).
Monensin impact on milk yield and milk components were extensively studied in dairy cows (Beckett et al., 1998; Duffield et al., 2000; Dubuc et al., 2010). No monensin effect on milk yield was detected in the present study. This is in agreement with Brown and Hogue (1985) results which, fed monensin to French Alpine lactating goats. Results on the effects of monensin on milk production in dairy cows are, however, inconsistent in the literature. Some studies have reported that monensin increased milk production, but others have not (Duffield et al., 2008). The increasing effect has been attributed to an increased supply of glucogenic precursors resulting from changes in the pattern of rumen fermentation (Phipps et al., 2000).
There is conflicting evidence on the effect of monensin supplementation on milk fat and protein. Monensin treatment decreased milk fat percentage by 1.25 percentage point and milk fat yield by 16 g/d (Table 5). This decreasing effect of monensin on milk fat percentage is in relative agreement with the results of meta-analysis by Duffield et al. (2008) in dairy cows. In addition, they found no effect of monensin on milk fat yield, which is similar to our results. The decreasing effect of monensin on milk fat could be explained by the fact that monensin causes a reduction of rumen biohydrogenation of long-chain fatty acids (Fellner et al., 1997), resulting in the production of specific fatty acid isomers (Bauman and Griinari, 2003) that inhibit milk fat synthesis in the mammary gland (Baumgard et al., 2000).
Monensin did not significantly alter milk protein yield and percentage in our study. These results were similar to those of Sauer et al. (1989), Beckett et al. (1998), and Duffield et al. (2000) in dairy cattle. No monensin supplementation effect was found on milk lactose percentage.
CONCLUSIONS
To the knowledge of authors this is the first comprehensive evaluation of the impact of monensin on metabolic parameters, dry matter intake and milk production in periparturient dairy goats. Monensin reduced serum BHBA concentration in the postpartum period. However, this improved energy metabolism did not result in increased milk production or improved dry matter intake. On the other hand, milk fat percentage significantly decreased in Monensin fed goats, which is not desirable quality for goat milk. Monensin significantly decreased DMI during the pre-partum period. However, Monensin is a promising tool for improving the energy status of dairy goats during transition period. Further work will be needed to document any potential health or performance benefits attributable to this improved energy supply.
Acknowledgements
This study was supported by research fund of Ferdowsi University of Mashhad (Project number: 364, 88/9/24).