Profiles of coagulase-positive and -negative staphylococci in retail pork: prevalence, antimicrobial resistance, enterotoxigenicity, and virulence factors
Article information

Abstract
Objective
The present study aimed to investigate the occurrence and species of coagulase-positive staphylococci (CoPS) and coagulase-negative staphylococci (CoNS) in retail pork meat samples collected during nationwide monitoring. The staphylococcal isolates were characterized for antimicrobial and zinc chloride resistance and enterotoxigenic potential.
Methods
A total of 260 pre-packaged pork meat samples were collected from 35 retail markets in 8 provinces in Korea for isolation of staphylococci. Antimicrobial and zinc chloride resistance phenotypes, and genes associated with the resistance phenotypes were determined on the isolates. Furthermore, the presence and distribution of 19 staphylococcal enterotoxin (SE) genes and enterotoxin-like genes among the pork-associated staphylococci were determined by multiplex polymerase chain reaction-based assays using the specific primer sets.
Results
A total of 29 staphylococcal strains (29/260, 11.1%) were isolated from samples of retail pork meat, 24 (83%) of which were CoNS. The four CoNS species identified were S. saprophyticus (n = 16, 55%), S. sciuri (n = 3, 10%), S. warneri (n = 3, 10%), and S. epidermidis (n = 2, 7%). Among the 29 isolates, four methicillin-resistant CoNS (MR-CoNS; three S. sciuri and one S. epidermidis) and one methicillin-resistant CoPS (MR-CoPS; one S. aureus) were identified. In addition, a relatively high level of tetracycline (TET) resistance (52%) was confirmed in CoNS, along with a predominant distribution of tet(K). The most prevalent SEs were sep (45%), and sen (28%), which were carried by 81% of S. saprophyticus.
Conclusion
These findings suggest that CoNS, especially S. saprophyticus strains, in raw pork meat could be a potential risk factor for staphylococcal food poisoning (SFP), and therefore, requires further investigation to elucidate the role of SEls in SFP and virulence of the pathogen. Our results also suggest that CoNS from raw pork meat may act as a source for transmission of antimicrobial resistance genes such as staphylococcal cassette chromosome mec and tet(K).
INTRODUCTION
Staphylococci are commensal colonizers of the skin and mucous membranes of humans and other warm-blooded animals. Based on their ability to coagulate rabbit plasma, staphylococci have traditionally been grouped into coagulase-positive staphylococci (CoPS) or coagulase-negative staphylococci (CoNS). As a major human and animal pathogen CoPS, S. aureus can cause a variety of infections including superficial skin and soft tissue infections, osteomyelitis, and septicemia [1]. Although not usually life-threatening, staphylococcal food poisoning (SFP) is one of the most frequent food-borne diseases, and results from the ingestion of staphylococcal enterotoxins (SEs) that are secreted mainly by CoPS, especially S. aureus [1]. SFP is a noncontagious acute gastrointestinal disease caused shortly after ingestion of food contaminated with enterotoxin-producing staphylococci. Frequency of SFP is often associated with economic impact such as loss in productivity, financial burden for food industry, and public healthcare system [2]. Although a detailed pathogenic role of SE-producing CoNS in SFP has not been elucidated, recent studies have demonstrated that CoNS can also produce SEs and could be a potential cause of food poisoning [3]. Furthermore, classical SEs (SEA-SEE) and toxic shock syndrome toxin-1 (TSST-1) genes have been detected in CoNS isolated from human and animal clinical samples [3,4].
In addition to enterotoxigenicity, the incidence of SFP cases, especially those involving antimicrobial resistant (AMR) staphylococci such as methicillin-resistant staphylococci (MRS), has been increasing in recent years [4]. Methicillin-susceptible staphylococci (MSS) become MRS via acquisition of a staphylococcal cassette chromosome mec (SCCmec) containing the mecA gene. An increase in the carriage rate of these SCCmec elements in CoPS and CoNS from various food sources has also been demonstrated in several recent studies [5]. Wendlandt et al [6] also suggested that CoNS and MSS isolated from livestock farm environments represent a significant reservoir for transmission of AMR genes to MRS such as livestock-associated methicillin-resistant S. aureus (LA-MRSA). Therefore, enterotoxigenic staphylococci with enhanced resistance to antimicrobial drugs in food, especially livestock products, would raise significant public health concerns.
In swine production chains in European countries and Korea, one of the most frequent MRSA clones are CC398 (ST398 and ST541) LA-MRSA, which seem to be associated with the overuse of antibiotics in pig farms [7,8]. In addition to LA-MRSA, recent publications also suggested that CoNS and MSS in pig farm environments play an important role in transmission of antimicrobial resistance genes through horizontal gene transfer mechanism [5,9]. These observations raise a concern for transmission of antimicrobial resistant CoPS and CoNS via pork meat. However, the prevalence of CoNS in retail pork meat, their antimicrobial resistance profiles, and their enterotoxigenic characteristics are not well established.
The present study aimed to investigate the occurrence and species of CoPS and CoNS in retail pork meat samples collected during nationwide monitoring. In addition, antimicrobial resistance profiles, carriage of major antimicrobial resistance genes, and prevalence and distribution of SE genes and enterotoxin-like genes were determined in the pork-associated staphylococci. Moreover, susceptibilities to zinc chloride, urease activity, hemolytic activity, and proteolytic activity were examined to characterize the virulence phenotypes of the isolates.
MATERIALS AND METHODS
Sample collection
A total of 260 pre-packaged pork meat samples were collected from 35 retail markets in 8 provinces in Korea from July 2017 to May 2018: Gyeonggi (58 samples), Gangwon (42 samples), Chungcheong (55 samples), Jeolla (54 samples), and Gyeongsang (51 samples). Four or five retail markets from each province were randomly selected based on regional distribution. At least 3 to 4 different raw pork products including pork belly, loin, leg, blade shoulder, tenderloin, and arm shoulder were collected from each retail market. All meat samples were stored below 4°C in a container with ice packs and transported to the laboratory for isolation of staphylococci within 24 h of purchase.
Isolation and identification of staphylococci
Isolation of staphylococci from pork meat samples was performed as described previously with minor modification [7,9]. Briefly, pork meat samples (25 g) were homogenized for 1 min in 225 mL buffered peptone water in a sterile stomacher bag (3M, St. Paul, MN, USA) using a stomacher (HUKO, Seoul, Korea). Next, 1 mL of the homogenized solution was added to 9 mL tryptic soy broth (TSB) supplemented with 10% NaCl and incubated at 37°C for 24 h. After pre-enrichment, 10 μL aliquots were streaked onto Baired-Parker agar (BPA) supplemented with egg yolk and potassium tellurite (Becton Dickinson [BD], Sparks, MD, USA) and incubated at 37°C for 24 to 48 h. Presumptive S. aureus with black colonies with a clear halo and/or staphylococcal colonies with gray/black color were all selected and up to 3 colonies of staphylococci from each sample were subcultured on BPA for further confirmation. Staphylococcal species identification was performed using both 16S RNA sequencing and the Vitek 2 system (bioMérieux, Marcy-l’Étoile, France). Genomic DNA samples of staphylococci were prepared using a Genemed kit (Seoul, Korea) according to the manufacturer’s recommendations. All the selected colonies were subjected to antimicrobial susceptibility assays and antimicrobial resistance gene detection assays.
Determination of antimicrobial susceptibility
Antimicrobial susceptibility tests were performed on all the study strains according to the guidelines of Clinical and Laboratory Standards Institute (CLSI, 2017). Briefly, minimum inhibitory concentrations (MICs) to oxacillin and tetracycline (TET) were determined using a broth micro-dilution method or a standard E-test (bioMérieux, France). Susceptibilities to ampicillin (10 μg), cefoxitin (30 μg), gentamicin (30 μg), ciprofloxacin (5 μg), chloramphenicol (30 μg), clindamycin (2 μg), erythromycin (15 μg), sulfamethoxazole-trimethoprim (23.73 to 1.25 μg), quinopristin-dalfopristin (15 μg), rifampin (5 μg), fusidic acid (50 μg), and mupirocin (200 μg) were determined using a disc diffusion method on Mueller-Hinton agar plates (BD, USA). All the antimicrobial discs were purchased from BD. In addition to the antimicrobial susceptibility assays, MICs to zinc chloride (ranging from 0.125 to 32 mM) were determined using a standard agar dilution method on Mueller-Hinton agar (pH-adjusted to 5.5) as previously described [10].
Detection of antimicrobial resistant genes and staphylococcal cassette chromosome mec typing
Staphylococcal strains displaying resistance to oxacillin, cefoxitin, and TET were examined for the presence of resistance genes. For methicillin-resistant strains, presence of mecA and SCCmec types were determined as previously described [11]. SCCmec types were determined according to the combination of chromosomal cassette recombinase (ccr) and methicillin resistance (mec) regions. TET resistant strains were tested for the presence of tet(K), tet(L), tet(M), tet(O), and tet(S) as previously described [12].
Analyses of staphylococcal virulence factors
Urease activity was measured for 24 h with shaking (200 rpm). Urease activity was scored and classified into four groups according to intensity of color change: none (yellow), weak (orange), moderate (pink), or strong (purple).
Hemolytic activity was examined on 5% sheep blood agar after 24 h growth at 37°C. Presence of alpha-, beta-, or gamma-hemolytic activity was determined as previously described [13].
Proteolytic activity was assessed on skim milk agar (SMA) plates containing 10% SMA solution and 1.5% Bacto agar (BD Difco, USA) [14]. Briefly, 10 μL of the bacterial inoculum was dropped onto SMA plates and proteolytic activity was determined based on zones of clearing at 4°C and 37°C.
Detection of staphylococcal enterotoxin genes
Carriage of SE genes in the 29 staphylococcal isolates was determined as previously described [2,15]. The presence of 5 classical SEs (sea, seb, sec, sed, and see) and 13 newer SEs (seg, seh, sei, sej, sek, sel, sem, sen, seo, sep, seq, ser, and seu) was examined using multiplex polymerase chain reaction (PCR) assays according to previous publications [15]. In addition to the SE genes, toxic shock syndrome toxin-1 (tst1 gene) was assessed using specific PCR primers [15]. Genomic DNA samples from reference S. aureus strains (N315, FRI472, MW2, FRI913, and COL) were used as positive controls for each multiplex PCR [15].
RESULTS
Profiles of pork-associated staphylococci
A total of 29 staphylococcal strains (29/260, 11.1%) were isolated from 260 pork meat samples collected in Korea over the 11-month study period (Table 1). The most frequent pork-associated staphylococci observed in this investigation were S. saprophyticus, comprising 55% of the staphylococci isolated from the samples. As shown in Table 1, 5 CoPS and 24 CoNS (17.2% vs 82.8%) were identified among the 29 pork-associated staphylococci. While only two species of CoPS (S. hyicus and S. aureus) were detected in the samples, four different species of CoNS (S. saprophyticus, S. sciuri, S. warneri, and S. epidermidis) were identified.

Profiles of Staphylococcus spp. isolated from 260 pork meat samples
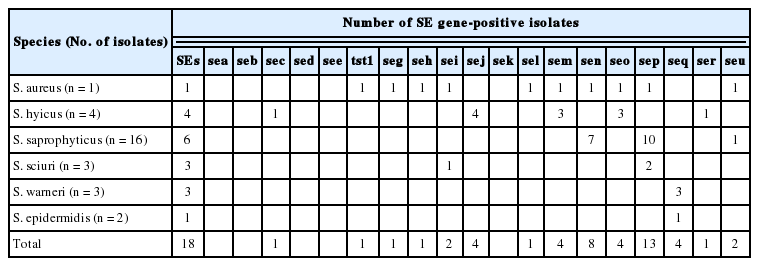
Carriage of SE genes in staphylococcal strains isolated from pork meat samples
The prevalence of staphylococci in the pork meat samples collected from different provinces is presented in Table 2. Interestingly, S. saprophyticus strains were isolated from all provinces, indicating that this is the most widespread staphylococcal species in pork meat samples in Korea. All the 29 staphylococci used in this study were isolated from different meat samples.
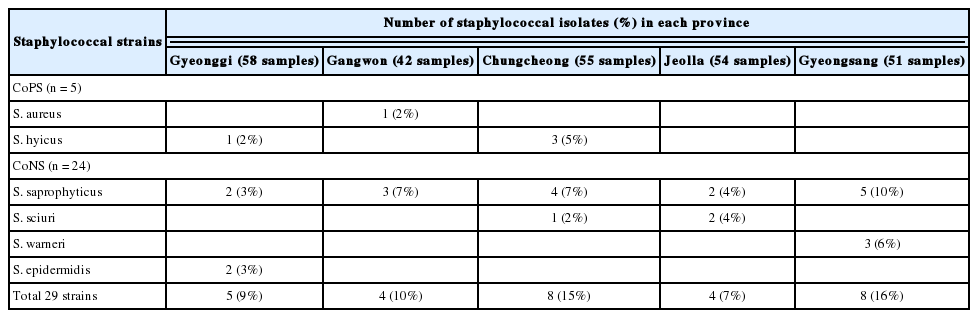
Prevalence of CoPS and CoNS in pork meat samples
Methicillin resistance among pork-associated staphylococci
Five MRS were identified among the 29 isolates (17%) collected from the pork meat samples (Table 1). These five isolates were all positive for mecA and displayed an OXA-resistant phenotype. SCCmec typing revealed that S. aureus (SA1), S. epidermidis (SE2), and S. sciuri (SS3) contained SCCmec II, SCCmec IV, and SCCmec V, respectively. Although two other S. sciuri strains (SS1 and SS2) were also positive for mecA, the SCCmec type remains undesignated because the ccr genes were not identified. The methicillin-resistant S. aureus strains were of sequence type (ST) 5, agr type II, and spa type t2460.
Antimicrobial resistance profiles of staphylococci isolates
All the 29 isolates were susceptible to chloramphenicol, sulfamethoxazole-trimethoprim, and rifampin (Table 3). A multidrug resistance (MDR) phenotype was observed in four staphylococci (SA1, SH4, SS1, and SE2), which showed resistance to ≥3 different classes of antimicrobials. The highest level of antimicrobial resistance was observed for TET (15/29 staphylococci, 52%), and all 15 TET-resistant staphylococci harbored one or two tet genes with TET MICs of ≥32 μg/mL. Of note, 14/15 TET-resistant isolates (93%) carried the tet(K) resistance gene.
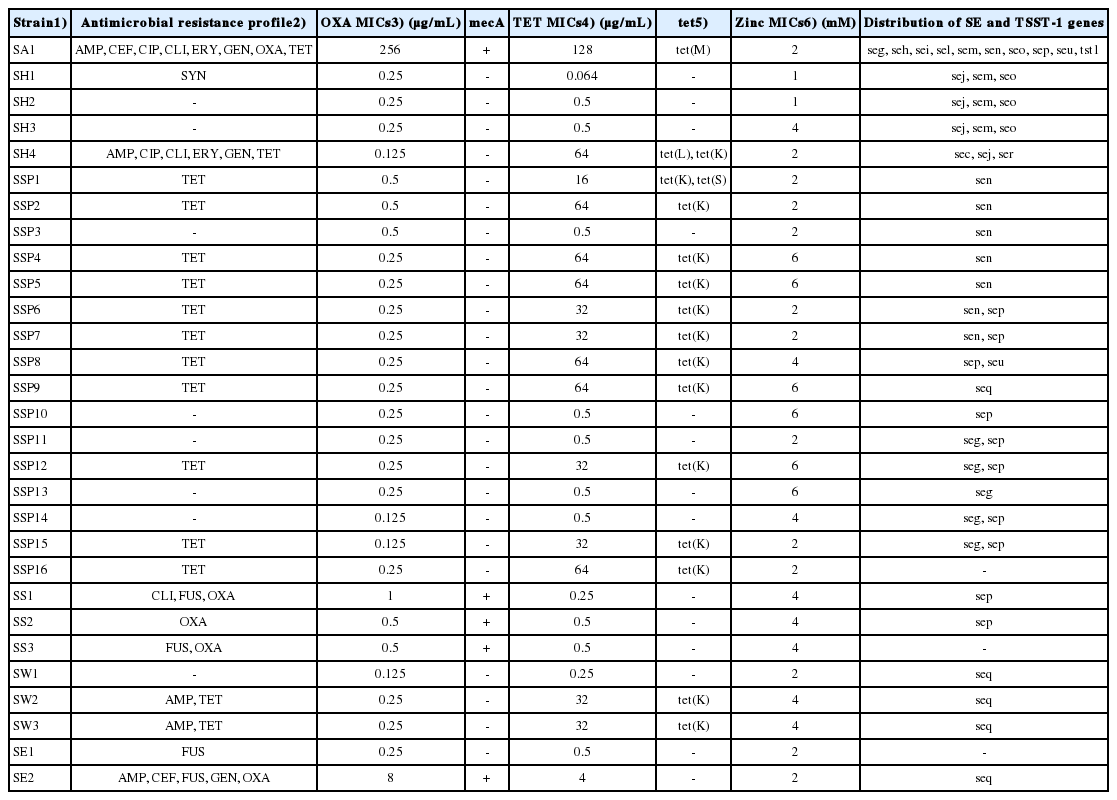
Antimicrobial resistance profiles, resistance genes, and zinc chloride resistance in CoPS and CoNS isolated from pork meat samples
Zinc chloride resistance has been proposed to be associated with methicillin resistance in specific clones of S. aureus of animal origin [7,16,17]. However, as shown in Table 3, 14/29 (48%) staphylococci displayed resistance to zinc chloride (MICs ≥2 mM) regardless of methicillin resistance.
Prevalence of staphylococcal enterotoxin genes
SEs are pyrogenic toxins and are involved in a significant number of human food poisoning diseases worldwide [2]. As shown in Table 4, all the staphylococcal isolates harbored one or more SE genes, except for three CoNS (SSP16, SS3, and SE1 strains), suggesting a widespread presence of SE genes among the CoPS and CoNS. A total of 11 different combinations of SE genes were identified among the 26 SE gene-positive staphylococci. Of the 18 SE genes analyzed, sep (13/29 isolates, 45%) and sen (8/29 isolates, 28%) were most frequently detected among the 29 staphylococci (Table 5). In particular, the sep and sen genes were present in 63% and 44% of S. saprophyticucs isolates, respectively. The SA1 strain (S. aureus) carried nine different SE genes and tst1, six of which are located within an enterotoxin gene cluster (seg, sei, sem, sen, seo, and seu). The presence of 6 enterotoxin genes (sen, sep, seu, seq, seg, and sec) identified in CoNS strains by PCR analyses were confirmed by sequencing PCR products (data not shown).
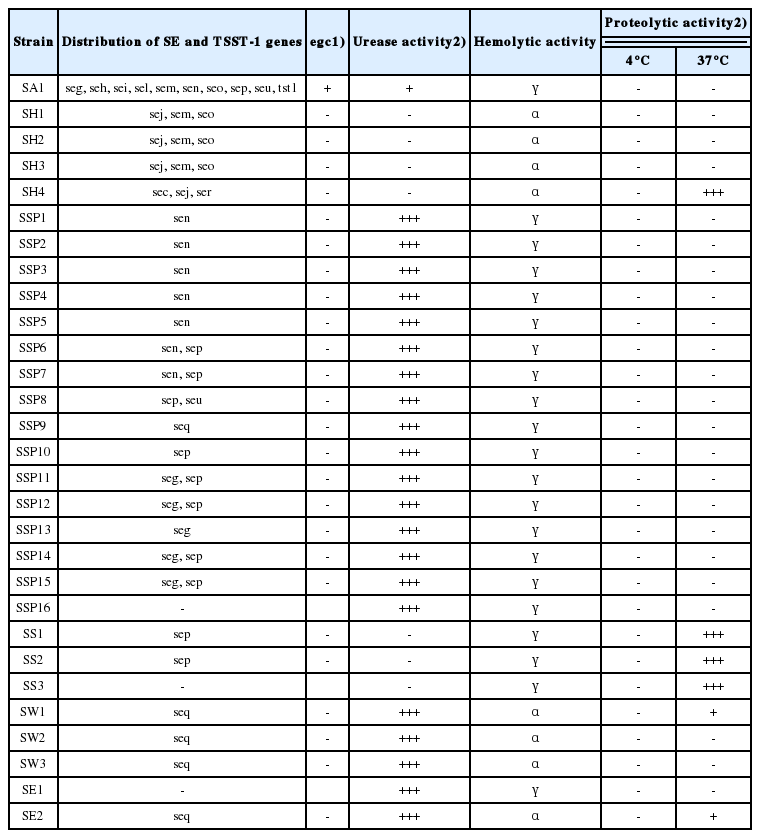
Carriage of SE and TSST-1 genes and exoenzyme activities in CoPS and CoNS isolated from pork meat samples
Assessment of virulence factors
Phenotypic analyses of hemolytic activity in the 29 staphylococci revealed that all isolates were hemolytic: three S. hyicus, three S. warneri, and one S. epidermidis were alpha-hemolytic and all others were gamma-hemolytic (Table 4).
Urease activity was mainly associated with CoNS, with an overall positive rate of 88%. Of note, 100% of S. saprophyticus, S. warneri, and S. epidermidis strains showed strong urease activity.
Strong proteolytic activity was observed only in one S. hyicus and three S. sciuri strains at 37°C. None of the 29 staphylococci displayed proteolytic activity at 4°C.
DISCUSSION
Although coagulase-positive S. aureus is the most common cause of SFP among staphylococcal species [1], recent studies have indicated that CoNS can produce SEs and become a potential cause of food poisoning. Staphylococci are normally present in the human and animal microbiota and are frequently found on the skin and in the upper respiratory tract of the host [18]. Thus, foods of animal origin such as milk, chicken, beef, and pork meats may represent a major source of SFP due to contamination with CoPS or CoNS during meat production. However, compared to S. aureus, only a few studies have addressed the prevalence and characteristics of non-S. aureus CoPS and CoNS in foods of animal origin.
In the present study, we investigated the prevalence of CoPS and CoNS in retail pork meat samples collected from July 2017 to May 2018 in Korea. The overall prevalence of staphylococci in the 260 pork meat samples was 11% (29 isolates/260 samples) and prevalence rates in five of the eight provinces ranged from 7% to 16%. Previous studies conducted in Nigeria, Poland, and Egypt reported prevalence rates of 40% to 91% for staphylococci in raw meats and meat products [4,19]. In particular, the prevalence of staphylococci in pork meat samples in Nigeria was 46%, which is significantly higher than the rate observed in the current study [19]. Consistent with prevalence rates reported in previous studies in Korea (0.4% to 19%), the prevalence of pork-associated S. aureus in the current study (0.4%, 1/260) was much lower than that reported for retail pork meat samples in the USA (16% to 67%) [20], Nigeria (54%), and China (18.6%) [21]. The present study reported that the proportion of CoNS observed among the 29 staphylococcal isolates was 83%, and this predominance of CoNS over CoPS is consistent with previous reports [4]. Four different CoNS species were identified and S. saprophyticus was detected with the highest incidence (16/29, 55%). In contrast, a predominance of S. epidermidis in raw pork meat was reported in a previous study from Nigeria [19]. These differences may have been caused by several factors such as geographical region, differences in pork production chains, and sampling/isolation methods. In addition, use of single methodology (enrichment in TSB supplemented with 10% NaCl and use of BPA) to isolate CoPS and CoNS would have affected the proportion of staphylococcal species presented in this study. However, these results suggest that CoNS isolates from raw pork meat could be a potential source of staphylococcal food contamination [4]. In addition, to the best of our knowledge, this is the first study to report the prevalence of CoPS as well as CoNS in retail pork meat collected nationwide in Korea.
MRS, especially methicillin-resistant S. aureus (MRSA), likely originated from CoNS by acquisition of SCCmec [22]. Methicillin-resistant CoNS have been isolated from a number of livestock and their products [3,4,23]. Consistent with these previous reports, four methicillin-resistant CoNS and one MRSA were isolated from raw pork meat samples in the current study. Although ST398 MRSA with SCCmec V has frequently been associated with livestock, particularly pigs [8], one strain of MRSA (SA1) isolated from raw pork was ST5 with SCCmec II. While one strain of S. epidermidis (SE2) and a strain of S. sciuri (SS3) was determined to harbor SCCmec IV and SCCmec V, two strains of mecA-positive S. sciuri (SS1 and SS2) could not be typed for SCCmec. Several studies have reported heterogeneity among SCCmec elements in MR-CoNS isolates [24]. In parallel with our results for S. sciuri, MR-S. sciuri carrying a non-typeable ccr gene was isolated from raw meat [25], indicating that a diverse pool of ccr genes and novel ccr types is present in S. sciuri isolates.
In addition to methicillin resistance, recent studies demonstrated that staphylococcal isolates from raw meat samples showed a TET resistance rate of 67% [26]. Similarly, a relatively high level of TET resistance (52%) was observed in staphylococci isolated in the current study, which is mainly attributed to TET-resistant S. saprophyticus isolates (11/16, 69%). TET resistance in staphylococci is mediated by acquisition of mobile tet genes, of which tet(K) has been most frequently associated with TET resistance in many CoNS of animal origin [3,18]. Consistent with previous observations [27], the present study found that 93% of TET-resistant staphylococci, and all the TET-resistant S. saprophyticus and S. sciuri isolates, carried tet(K).
A zinc resistance phenotype mediated by czrC within the SCCmec V element is typically associated with a high incidence of swine associated ST398 MRSA in European countries [10,28]. Recently, czrC-mediated zinc-resistance has also been reported in S. aureus strains isolated from meat products [29,30]. Unlike MRSA, data on the zinc resistance in non-aureus staphylococci (NAS) or CoNS are limited despite the possible co-selection of MRS using zinc in pig farms. However, the zinc-resistant gene czrC was identified in methicillin-resistant S. hyicus strains [31]. Moreover, frequent presence of czrC in methicillin resistant CoNS strains, such as S. epidermidis, S. haemolyticus, S. hominis, and S. lentus has been reported regardless of their SCCmec type [32]. Similarly, in the current study, 14/29 staphylococci displayed resistance to zinc chloride regardless of their methicillin or TET resistance, suggesting that co-selection of zinc resistance did not play a significant role in the high prevalence of methicillin- and TET-resistant CoNS strains used in this study. To the best of our knowledge, this is the first study in Korea that investigated the zinc resistance in NAS strains isolated from pork meat samples. Further studies are necessary to determine whether the use of zinc will be a major factor for co-selection of zinc and antimicrobial resistance in NAS or CoNS.
SEs were originally identified in S. aureus and are known to be responsible for SFP in humans. In addition to the 5 classical SE genes (sea, seb, sec, sed, and see), 19 newer SEs, and enterotoxin-like (SEl) peptides have been identified [2]. These SEs and SEls are mainly associated with CoPS S. aureus-associated SFP worldwide [2]. However, recent studies reported CoNS such as S. epidermidis, S. chromogenes, S. hyicus, and S. haemolyticus isolates of human and animal origin carried SE genes [4,15]. In the present study, we found that CoNS isolated from pork meat samples more often possessed the newly described SEs and SEls rather than the classical SEs. Moreover, our data showed that sep and sen were the most common SEs among the CoNS, especially in S. saprophyticus isolates. In contrast to the high prevalence of sep and sen in our CoNS, previous studies reported that sec was the most prevalent SE gene in CoNS isolates [33]. More recently, three classical SEs (sea, seb, and sec) were identified as the most frequent enterotoxins among CoNS isolated from bovine milk [23]. Although the role of SEls in outbreaks of SFP is still controversial, the predominant presence of enterotoxigenic CoNS in raw pork meat could represent a potential food safety hazard. The SEs are synthesized and secreted by staphylococci throughout the exponential growth phase and during the transition from exponential to stationary growth phases [34,35]. Usually, the infective dose of enterotoxins needed to induce SFP in humans is only high nanogram to low microgram ranges. Synthesis of SEs is regulated by multiple factors such as bacterial cell density, changes in microenvironment, harsh growth conditions, and changes in bacterial cell membrane physiology [2,34,35]. These factors affect expression of enterotoxins through the alternative sigma factor (σB), SarA, Agr quorum-sensing system, and SaeRS two-component regulatory system [2,34]. Unlike the classical SEs, information on the regulation of newer enterotoxins is limited even in coagulase-positive S. aureus. Therefore, further studies are necessary to characterize expression profiles of enterotoxin genes found in the CoNS, especially sep and sen genes in S. saprophyticus strains.
Staphylococci can produce secretary proteins such as pro teases and lipases that may have a negative effect on food preservation, causing a deterioration in quality and food spoilage [36]. Although protease-induced food spoilage has typically been associated with Pseudomonas spp. and psychrotrophic bacteria [36], pork meat derived coagulase-negative S. hyicus and S. sciuri strains showed strong protease activity at 37°C. In addition, strong urease activity, which is often associated with acute urinary tract infections, was only observed in CoNS species of S. saprophyticus, S. warneri, and S. epidermidis.
It should be recognized that the current study has several limitations. Our results were generated from a rather limited number of samples and staphylococcal isolates, which warrants future investigations with a larger number of various meat samples. In addition, complete genotypic characterization of the CoPS and CoNS for antimicrobial resistance, virulence, and synthesis of SEs in culture supernatants was not included in the current investigation.
CONCLUSION
Our results demonstrate the following: i) a relatively high level of CoNS, especially S. saprophyticus species, are present in retail pork meat; ii) while the occurrence of MDR in pork-associated CoNS was low, a high level of TET resistance among CoNS was observed, along with a predominant possession of tet(K); and iii) a frequent presence of newly defined SEls such as sep and sen were confirmed in CoNS species, especially in S. saprophyticus strains isolated from pork meat. Thus, in addition to CoPS, our results suggest that CoNS from raw pork meat could serve as a potential reservoir for antimicrobial resistance as well as enterotoxins typically associated with SFP. Future studies are also necessary to investigate prevalence and characteristics of CoPS and CoNS in other foods of animal origin such as milk, chicken, and beef meats.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.
ACKNOWLEDGMENTS
This research was supported by funding from the Research of Korea Centers for Disease Control and Prevention (2020 ER540500 to S.J.Y.).
We thank Haeng Ho Lee and Hong Sik Eom for excellent technical assistance with isolation of staphylococci and antimicrobial susceptibility assay.