Immunomodulatory effects of phytogenics in chickens and pigs — A review
Article information

Abstract
Environmental stressors like pathogens and toxins may depress the animal immune system through invasion of the gastrointestinal tract (GIT) tract, where they may impair performance and production, as well as lead to increased mortality rates. Therefore, protection of the GIT tract and improving animal health are top priorities in animal production. Being natural-sourced materials, phytochemicals are potential feed additives possessing multiple functions, including: anti-inflammatory, anti-fungal, anti-viral and antioxidative properties. This paper focuses on immunity-related physiological parameters regulated by phytochemicals, such as carvacrol, cinnamaldehyde, curcumin, and thymol; many studies have proven that these phytochemicals can improve animal performance and production. On the molecular level, the impact of inflammatory gene expression on underlying mechanisms was also examined, as were the effects of environmental stimuli and phytochemicals in initiating nuclear factor kappa B and mitogen-activated protein kinases signaling pathways and improving health conditions.
INTRODUCTION
Pathogens like viruses, bacteria, fungi and other environmental stimuli are latent stressors which may damage animal tissues and cells. Excessive stressors may increase the concentration of reactive oxygen species (ROS), causing lipid peroxidation and oxidative damage to cellular membranes. The lack of sufficient antioxidants to eliminate ROS will lead to oxidative damage and trigger inflammation [1–3]. When inflammation occurs, cells have to recruit other cells to local sites by secreting more inflammatory cytokines and chemokines; this is a necessary process for the host to fight against infection and to repair tissue. However, prolonged inflammation may lead to unnecessary energy expenditure. As such, the alleviation and prevention of over-inflammation and the return of immune status to normal condition will benefit animal production.
As the first line of defense, the gastrointestinal tract (GIT) has a large surface area where food degradation and absorption take place; pathogens and toxins may also gain access to the body through the GIT by destroying the mucus layer [4]. Accordingly, protection of the GIT against the incursion of pathogens and toxins is of vital importance. Common toxins, such as aflatoxin and lipopolysaccharide (LPS), may cause great damage to intestinal cells, induce inflammation and immune response, and impair production [5,6]. Toxic airborne gases, like ammonia, may harm the respiratory system of animals, resulting in depressed performance [7,8]. Exogenous stressors may exacerbate the pathogenic invasion in the gut of animals, indicated by decreased bodyweight and feed intake, infiltration of heterophils in the intestine and increased Salmonella numbers in the spleens of heat stressed chickens after Salmonella infection [9]. Coccidiosis, a parasitic disease in birds infected mainly with Eimeria spp., is a major problem in the poultry industries worldwide, resulting in enormous annual losses [10]. These stressors impair mucosal immune responses to other pathogens and cause intestinal damage and poor growth performance [6,11]. Therefore, many different feed additives, like antibiotics, probiotics, prebiotics and phytogenics, have been added to animal diets to improve the hosts’ capacity to eliminate the negative influence of environmental stressors and improve growth performance [12–17].
Antibiotics had been widely used in the livestock industry, but adverse effects, including drug resistance and reduced beneficial endogenous bacteria counts, led to subsequent prohibition against antibiotic supplementation by the European Union in 2006. Many other governments also began to restrict antibiotic supplementation, prompting a great deal of new research into phytochemicals as alternative feed additives. It was observed that supplementation of phytochemicals had positive effects on the pancreatic and intestinal enzyme secretion in broilers [18,19], and improved dry matter and crude protein digestibility in pigs [20]. Although inconsistent results of feed intake suggested that phytogenics might not improve feed palatability, feeding phytogenics did promote nutrient absorption and feed conversion ratio (FCR) in poultry [21]. Many authors have pointed out that phytogenics in animal diet improved the immune response and the protective capacity of the GIT against exogenous stressors [22–26]; however, the mechanisms by which phytochemicals act on host cells were still unclear. In this review, we summarize the known relevant mechanisms in mammals and poultry, and suggest the potential immunomodulatory effects of phytochemicals on chickens and pigs.
EFFECTS OF PHYTOCHMICALS ON IMMUNE RESPONSE
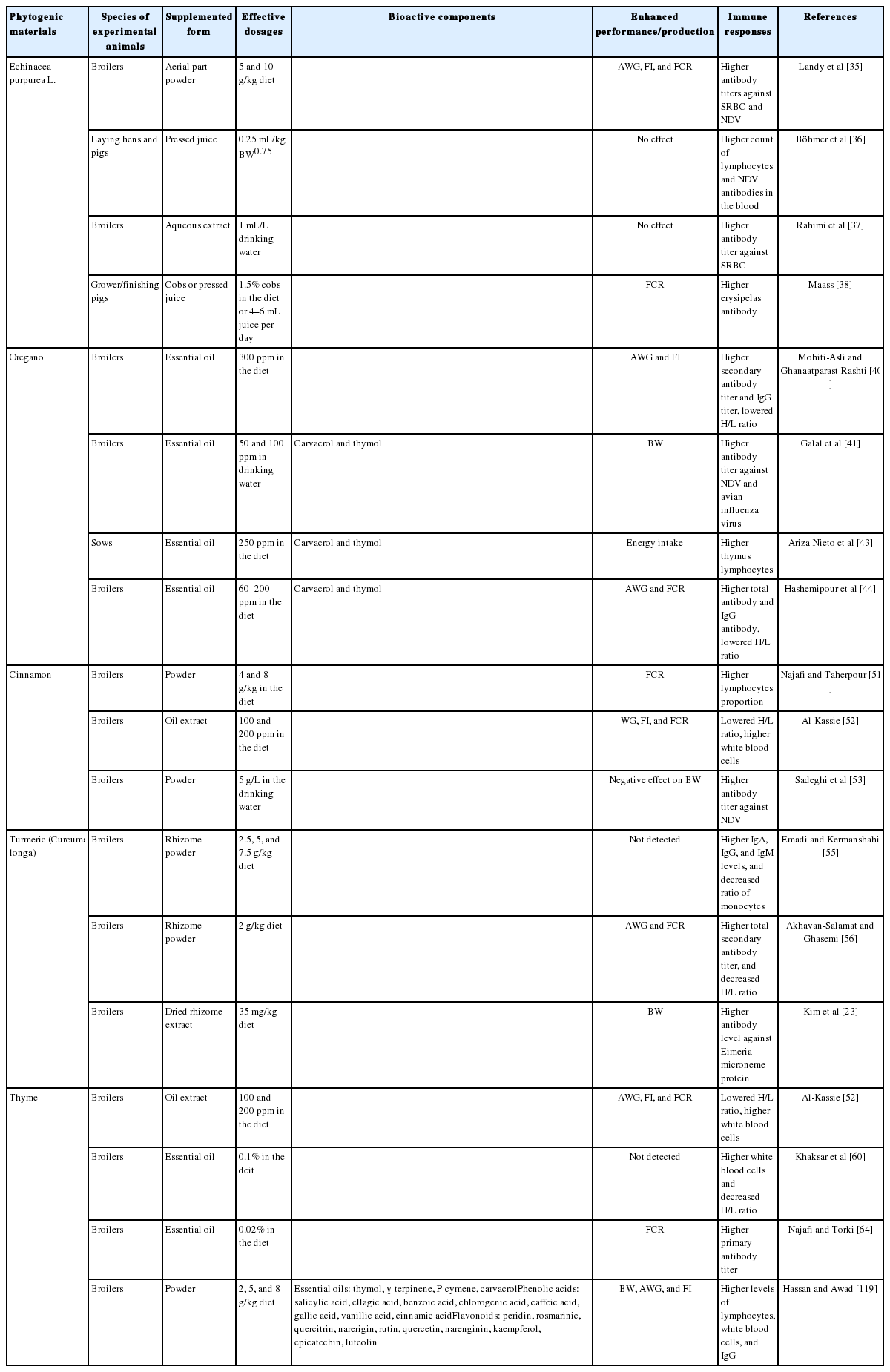
Phytochemicals, the active constituents in phytogenics, can be roughly divided into two groups based on the different synthetic pathways in plants: terpenes and terpenoids, or aromatic and aliphatic compounds [27]. Some phenolic compounds and terpenes have been reported to exhibit anti-inflammatory activity [28,29]. However, as all the phytochemicals were extracted under different conditions or by using different parts of the plant, it is difficult to evaluate their effect. Phytogenics have traditionally been used in powder form, but antagonizing or additive reactions between compounds in a single phytogenic may result, as inconsistent results of immunoregulatory action between treatments of phytogenic and its individual component were found [30,31]. It’s difficult to study the possible interactions between compounds in phytogenics, not to mention the diversity of phytogenics and phytochemicals. As such, the complexity of the immunoregulatory actions of phytogenics limits the progress of related research. Nevertheless, phytogenics and phytochemicals still remain among the most promising feed additives thanks to their positive effect on regulating the immune response in chickens and pigs. Here we review the effects of some popularly applied phytogenics on the immune response in animals, including Echinacea purpurea L. (EP), oregano, cinnamon, Curcuma longa, and thyme (Table 1).
Effect of five phytogenics, Echinacea purpurea L., oregano, cinnamon, turmeric, and thyme, on the immune responses of chickens and pigs
Echinacea purpurea L
In our previous studies, EP extract, the herbal medicine normally used to prevent respiratory infections in humans, was evaluated for its antioxidant activity in broilers [32–35]. The results demonstrated that EP ethanol extracts had no major influence on the viability of the mononuclear cells in peripheral blood [32]. Dietary supplementation with dried EP improved both the FCR and meat quality [33,34]. It was also reported that 5.0 g/kg EP supplementation in broilers could increase the antibody titer against sheep red blood cells (SRBCs) and Newcastle disease virus (NDV) [35]. The EP, which was fed to laying hens and fattening pigs as pressed juice preserved with ethanol, still showed immunomodulatory effects, as indicated by the increased count of lymphocytes and NDV antibodies in the blood [36]. The 0.1% supplementation of EP aqueous extract in the diet enhanced cutaneous basophil hypersensitivity and antibody response against SRBCs in broilers [37]. However, no significant differences were observed in sows fed 1.2% and 3.6% EP cobs during gestation, in sows fed 0.5% and 1.5% cobs during lactation, or in piglets fed 1.8% cobs compared to the control group, while the FCR and erysipelas antibodies were improved in grower-finisher pigs supplemented with 1.5% cobs or 4 to 6 mL pressed juice per day [38]. Accordingly, EP supplementation may promote host immunity in broilers and fattening pigs, but determining its immunomodulatory effect and effective dosage in sows and piglets needs more research.
Oregano
Oregano belongs to the Labiatae family. Oregano is the popular name for a variety of widely distributed plant species, including Origanum vulgare L. and O. onites L., and is characterized as containing great amounts of carvacrol as the main source of its fragrance. Another plant called oregano is Coridothymus capitatus (L.) Reichenb. fil. (Spanish oregano) [39]; it has been widely studied as a feed additive in animals. Mohiti-Asli and Ghanaatparast-Rashti [40] reported that dietary supplementation with 300 ppm of oregano essential oil (OEO) improved the European production efficiency factor in broilers, which was calculated according to body weight, livability, FCR and trial duration; 500 ppm OEO had no positive effect, while 300 ppm OEO in the diet produced higher secondary total antibody titer against SRBCs; in addition, the immunoglobulin G (IgG) titer was higher than it was in the control group. Another study reported that higher doses (500 and 1,000 ppm) of OEO supplementation led to positive immune responses in NDV- and avian influenza-vaccinated broilers [41]. Serum heterophil counts and the heterophil to lymphocyte ratio (H/L ratio), both considered stress indicators [42], were decreased in birds fed 300 ppm OEO. On day 14 of lactation, higher thymus lymphocyte concentrations were observed in sows’ milk after 250 ppm OEO was added to their diet, indicating the immunostimulatory effect of OEO [43]. Other studies also reported that 300 ppm OEO or 300 ppm of a mixture of carvacrol and thymol could improve the growth performance or the immune response, as indicated by lowered excreta oocyst counts of coccidia [24], and increased hypersensitivity response, total antibody titer, and H/L ratio [44]. However, the growth promoting effects of OEO were inconsistent [45], probably because the immunomodulatory actions of OEO could not improve growth performance in a less harsh experimental environment when compared to practical rearing conditions.
Cinnamon
The essential oils from some Cinnamomum species contain great amounts of trans-cinnamaldehyde (C. burmannii), while other species do not (C. tamala, C. pauciflorum, and C. zeylanicum) [46,47]. Dietary supplementation with cinnamon leaves or extracted oil have been demonstrated to improve growth performance in broilers [48,49], and decrease coliform bacteria in the jejunum and large intestine [50]. Najafi and Taherpour [51] reported that the inclusion of 0.4% and 0.8% cinnamon in broiler diets improved the FCR and increased the concentration of hemoglobin and the proportion of lymphocytes in the blood. Dosages of 100 and 200 ppm essential cinnamon oil improved FCR and body weight gain from 1 to 6 weeks, reduced the H/L ratio and increased the level of white blood cells in broilers [52]. Conversely, positively modulated immune responses and impaired growth performance were detected in 21-day-old broilers that were given 5.0 g/L cinnamon in their drinking water [53]. This discrepancy may be because the majority of studies supplied cinnamon to animals via their diet versus their drinking water. Furthermore, the variety of species used in the different experiments make it difficult to determine the effective dosage of cinnamon or cinnamon oil, or to make valid comparisons across studies, especially if the content of the major bioactive compounds was not measured.
Turmeric (Curcuma longa)
There are many species in the genus Curcuma, among which C. longa (turmeric) is the most studied. The major phytochemicals in the rhizome of C. longa vary dramatically with the environment and location. Li et al [54] concluded that curcumin, demethoxycurcumin and bisdemethoxycurcumin are the main compounds in the rhizomes, powders and extract products, whereas the concentrations of ar-turmerone, α-turmerone, and β-turmerone are related to the quality of the turmeric oils and oleoresin products. Most studies evaluated this phytogenic in the form of turmeric rhizome powder (TRP). TRP inclusion in the diet significantly increased blood IgA, IgG, and IgM levels, and decreased the ratio of monocytes in 42-day-old broilers challenged with SRBCs [55]. Similarly, 0.2% TRP supplementation decreased the H/L ratio and increased the total secondary antibody titer against SRBCs in broilers undergoing heat stress treatment [56]. Moreover, the addition of 0.33% and 1.0% TRP, which contained 1.16% curcumin, improved the FCR in broilers during the entire feeding period [57]. Inclusions of 0.33%, 0.66%, and 1.0% TRP significantly reduced abdominal fat content and serum triglyceride concentration [57]. Therefore, dietary curcumin could regulate immune responses in animals; it has the potential to enhance growth performance.
Thyme
Thyme is the popular name for Thymus spp.; thymol, carvacrol, p-cymene and γ-terpinene are the major active components in the essential oil [58,59]. Dietary supplementation of thyme was in the form of essential oil or leaf powder. It has been shown that supplementation with thyme essential oil improved cutaneous basophil hypersensitivity to phytohaemagglutinin-P (PHA-P) and reduced the H/L ratio. It also produced a more balanced intestinal microbiology, as indicated by higher Lactobacillus and Bifidobacterium counts and lower Escherichia coli (E. coli) levels in the ileum [60]. Previous studies suggested that heat stress may impair the cell-mediated immune response to PHA-P in chicks [61], highlighting the benefit of feeding thyme essential oil to birds. The H/L ratio is a stress indicator, as mentioned above. A reduced H/L ratio demonstrates the enhanced immunoregulatory capacity of the host. Intestinal microbiota plays an important role in the immune system of chickens [62]. Environmental stressors may change the intestinal microbiota and increase the susceptibility to colonization by pathogens in the intestine [63]. It was also reported that thyme essential oil improved bodyweight gain and FCR in broilers [52,64]. As such, thyme has potential as a feed additive in poultry production.
INTERACTIONS BETWEEN PHYTOCHEMICALS AND THE MAPK- AND NFκB-REGULATED PATHWAYS IN CHICKENS AND PIGS
NFκB and MAPKs in immune responses
Innate immunity, also known as non-specific immunity, includes activation and the recruitment of innate leukocytes like macrophages, thereby enhancing phagocytosis. As pathogens invade, innate leukocytes identify and engulf them while expressing cytokines, thereby initiating the immune response and leading to an inflammatory reaction [65]. Adaptive immunity is a specific immune system process initiated by the presentation of antigens. Pathogens and exogenous stimuli are identified and eliminated by B cells and T cells. Among these, helper T cells are induced by effector cytokines to differentiate into cell types Th1 or Th2, and subsequently deliver signals to macrophages or B cells, respectively [66]. Therefore, homeostasis is maintained through the mutual actions of the innate and adaptive immune systems.
In the innate immune system, toll-like receptors (TLRs) are the most studied pattern-recognition receptors in poultry and livestock [67–69]. As TLRs are stimulated, many signaling pathways are activated simultaneously through adaptor proteins such as myeloid differentiation primary response 88 (MyD88) and toll-IL-1-resistance domain-containing adaptor, inducing interferon β [70]. The Src kinase plays a vital role in the activation of mitogen-activated protein kinases (MAPKs) and nuclear factor kappa B (NFκB), the two major pathways that modulate immune responses [71–73]. MAPKs are a group of protein kinases that regulate cellular activities. Among MAPKs, p38 protein kinase, extracellular signal-regulated protein kinase (ERK1/2), and c-Jun N-terminal kinase have been reported to be associated with the regulation of inflammation and immunity in response to environmental stress [74–76]. The NFκB-mediated pathway involves many physical reactions and is extremely important in the modulation of immunity, inflammation and apoptosis [71]. It is well known that NFκB is a dimer consisting of different subunits [71]. Under normal physical conditions, the dimer binds to the inhibitor of kappa B (IκB) and presents as an inactive form in the cytosol. When the cell is activated by exogenous stimuli, NFκB is liberated from its IκB-inhibited form. The inhibitor of kappa B kinase (IKK) phosphorylates the linked IκB, leading to the ubiquitination of phosphorylated IκB, and the subsequent identification and degradation by the 26S proteasome [77]. Disconnected NFκB enters the nucleus, binding to a specific region of DNA, and promotes the expression of downstream genes. Translated proteins are secreted into the extracellular space to modulate the immune system and inflammatory response [78]. Depending on the strength, timing and site of stimulation, different downstream products are generated and exert diverse functions, including cellular growth, differentiation, tissue repair and synthesis of tissue collagen. Therefore, studies on NFκB signaling include its onset and therapies for many diseases as well as the apoptosis of cancer cells [79]. Pro-inflammatory cytokines, ROS and mitogens generated by bacterial or viral infection, chemical and biological tissue damage or other stressors, induce the activation of NFκB signaling [80,81], followed by the expression of more inflammatory cytokines and chemokines. Interleukin 1 (IL-1), IL-6, and tumor necrosis factor alpha (TNF-α) are the three most investigated pro-inflammatory cytokines that can be expressed by monocytes and macrophages after invading pathogens are identified. These cytokines mediate metabolic changes, which enhance the immune response and disease resistance, thereby stunting growth performance [82,83]. TNF-α may be secreted by activated macrophages and lead to chemotaxis of monocytes [84]. Expressions of IL-1β, IL-6 and TNF-α are highly associated with the immune status of poultry and livestock [85].
Nitric oxide (NO) and prostaglandin E2 (PGE2) are important inflammatory mediators that modulate the development of inflammation [86,87]. After NFκB is activated, NFκB-mediated signaling molecules upregulate the expression of the downstream gene COX-2, synthesizing cyclooxygenase 2, which converts arachidonic acid into PGE2. Inducible nitric oxide synthase (iNOS) is also activated, which subsequently increases NO production [88]. The enhanced production of histamine and PGE2 in response to stressors not only induces phagocytosis of macrophages, but also affects the production of cytokines by T cells, macrophages and dendritic cells [87,89]. These cytokines include IL-1 and IL-6, which are transported through the circulatory system to the brain, activating the hypothalamic-pituitary-adrenal (HPA) axis [90]. In response to acute inflammation, the HPA axis is stimulated to secrete more glucocorticoids, which have been thought to elevate the H/L ratio in birds and the neutrophil to lymphocyte ratio in mammals. Glucocorticoids also induce heterophils (neutrophils) to express pro-inflammatory cytokines, including IL-1β, IL-6, IL-10, IL-12α, and IL-18 [91]. Increased heterophils (neutrophils) and decreased lymphocytes in the circulatory system represent the development of the innate response in the host as inflammation is triggered. Elevated H/L ratio is considered a stress marker. Exogenous substances are eliminated through acute inflammation in organisms. However, under stressful conditions, the suppressed immune system is unable to cope, leading to chronic inflammation [92]. It has been proven that chronic inflammation is associated with bacterial disease and heat stress in animals [9,93,94]. In the previous section, we reviewed that phytogenics, such as oregano, cinnamon, turmeric, and thyme, are able to modulate humoral responses in chickens and pigs. Their major components have been reported to regulate NFκB and/or MAPKs signaling pathways in rodents [95–97]. Therefore, phytochemicals that modulate the immune response and regulate the expression of inflammatory mediators in chickens and pigs were introduced (Figure 1); they are listed below.

The possible immunomodulatory action of carvacrol, cinnamaldehyde, curcumin, and thymol regulating expression of pro-inflammatory mediators and enzymes involved in the nuclear factor kappa B (NFκB) and mitogen-activated protein kinases (MAPKs) signaling pathways, inflammation, and immune response in chickens and pigs.
Carvacrol
Carvacrol (2-methyl-5-(1-methylethyl)-phenol), the major active component in oregano and thyme essential oils, is a phenolic monoterpene. In mice, carvacrol reduced local levels of IL-1β and PGE2, and mRNA levels of COX-2 and IL-1β [98]. The mRNA and protein levels of TNF-α, IL-6, Inos, and COX-2 were also downregulated in D-galactosamine-induced hepatotoxic rats after treatment with carvacrol. The suppression of pro-inflammatory cytokine production may be attributed to the inhibition of TLR2- and TLR4-mediated signaling [99]. Zou et al [100] reported that OEO, which contains 81.92% carvacrol, lowered the protein level of MAPKs and NFκB, and decreased the expression of the pro-inflammatory cytokines IL-1β, IL-6, interferon-γ (IFN-γ), TNF-α and monocyte chemotactic protein-1 in the jejuna of pigs, proving its anti-inflammatory effects. Moreover, the mRNA levels of occludin and zonula occludens-1, two epithelial tight junction proteins, were increased, demonstrating that carvacrol may improve the integrity of intestinal epithelia. Reduced expression of IL-6 and LPS-induced TNF-α factor were observed in Eimeria oocyst-challenged broilers supplemented with essential oils containing 81.89% carvacrol [101]. It is speculated that carvacrol may suppress TLRs and/or MAPKs and NFκB signaling pathways in animals; however, more studies are needed to elucidate its mechanisms in detail.
Cinnamaldehyde
In vitro studies have demonstrated that both Cinnamomum cassia oil and cinnamaldehyde, the major bioactive compound in cinnamon, inhibited mRNA expression of pro-inflammatory cytokines, including IL-1β, IL-6, and TNF-α, and increased the production of anti-inflammatory cytokine IL-10 in LPS-activated J774A.1 cells. Subsequently, the expression of iNOS and COX-2 was decreased, as was NO production [102]. Youn et al [95] reported that cinnamaldehyde could suppress LPS-induced activation of NFκB and interferon regulatory factor 3, resulting in the decreased expression of the TLR4 target genes COX-2 and IFN-β. In broilers, an analysis of global gene expression profiles indicated that the immunomodulatory effects of dietary cinnamaldehyde were related to antigen presentation, humoral immune response and inflammatory disease [25]. In vitro tests showed that chicken spleen lymphocytes treated with cinnamaldehyde (25 to 400 ng/mL) significantly promoted cell proliferation compared to the control group. The dosages of 1.2 to 5.0 μg/mL of cinnamaldehyde stimulated macrophages to produce more NO, while 0.6 to 2.5 μg/mL inhibited the growth of chicken tumor cells; 10 and 100 μg/mL of cinnamaldehyde lowered the viability of Eimeria tenella [103]. Conversely, in vivo test results showed that dietary supplementation with 14.4 mg/kg cinnamaldehyde increased mRNA expression of the pro-inflammatory cytokines IL-1β, IL-6, IL-15 and IFN-γ. Cinnamaldehyde supplementation also improved the body weight gain of E. acervulina or E. maxima-infected chickens [103]. It should be noted that the dosage of cinnamaldehyde used in this study was very low, and treatment with low concentrations of cinnamaldehyde (0.01 to 0.5 μg/mL) increased NFκB activation in LPS-stimulated human cells in vitro [104]. Further experiments are needed to prove whether cinnamaldehyde has a dose-dependent immunoregulatory effect on birds.
Curcumin
Curcumin (diferuloylmethane), one of the major components in turmeric, has been extensively researched. Previous studies have shown that curcumin involved in the p38 MAPK signaling pathway in LPS-stimulated RAW 264.7 macrophages [96], produced similar results in dendritic cells [105], and in the colonic tissue of rats with intracolonic injuries [106]. Additionally, Lubbad et al [107] reported that curcumin could suppress inflammation through the reduction of protein levels of TLR4, MyD88, and NFκB. MyD88 is normally recruited after TLR4 activation, and leads to the recruitment of more downstream molecules, as well as the activation of NFκB [108]. Acute carbon tetrachloride-induced liver damage was also alleviated by reducing the protein production of TNF-α and IL-1β in rats with curcumin treatment [109]. In poultry, the mRNA expression of IL-1β, IL-6, IL-12, IL-18 and tumor necrosis factor superfamily 15 were enhanced in chicken macrophages treated in vitro with organic extract of turmeric [110]. The mRNA expression of IL-6 in the liver of broilers was increased by aflatoxin B1 treatment, but alleviated by turmeric powder supplementation [111]. The results of microarray hybridization demonstrated that the gene expression of myeloperoxidase, cluster of differentiation 28 and lactotransferrin are associated with the inflammatory response in broilers [112–114], which was downregulated by treatment with organic extract of turmeric [23]. A series of studies was conducted to evaluate the effect of turmeric oleoresin, which contained 98% curcuminoids, on the immune response of swine [26,76, 115,116]. The supplementation significantly reduced serum IL-1β and TNF-α levels in porcine reproductive and respiratory syndrome virus-challenged pigs [26] and counteracted the effects of E. coli, as indicated by the markedly altered mRNA expression of IL-1β, mucin 2, COX-2 and p38 MAPK in ileal mucosa [76] and serum TNF-α concentration [115]. According to the above reports, in animals, curcumin mainly modulated the p38 MAPK signaling pathway and regulated downstream pro-inflammatory mediators, such as IL-1β, IL-6, and TNF-α.
Thymol
As the major compound in the essential oils of thyme and oregano, thymol is the isomer of carvacrol; thymol has been shown to possess anti-inflammatory properties; it suppresses the maturation of dendritic cells and activation of T cell proliferation in vitro [117]. Mature dendritic cells play an important role in the promotion of the immune response. Additionally, thymol inhibited the phosphorylation of NFκB and MAPKs, and lowered the production of IL-6, TNF-α, iNOS, and COX-2 in LPS-stimulated mouse mammary epithelial cells [97]. It also reduced the production of NO and hydrogen peroxide in LPS-stimulated cells [118]. In the jejuna of broilers, supplementation with thyme powder, which contains the essential oils thymol (50.48%), γ-terpinene (11.03%), P-cymene (9.77%), and carvacrol (4.30%), significantly decreased transcriptional factor NFκB. Pro-inflammatory cytokines such as IL-6, IFN-γ, and TNF-α were also downregulated in broilers following treatment with thyme powder [119]. This is consistent with previously mentioned results, suggesting that the anti-inflammatory effects of thyme make it suitable for application in animal production.
CONCLUSION
As it is inevitable for animals to be faced with environmental stressors like heat, disease and microbial toxins, the defense system of the host plays a vital role in maintaining health as well as growth performance and production. According to the above reports, the immune response improved when animal diets were supplemented with cinnamon, oregano, turmeric or thyme products. The stress response was alleviated since the NFκB and MAPKs signaling pathways were suppressed, and the expression of anti-inflammatory cytokines was increased. As a result, the blood levels of non-specific immune cells such as heterophils (neutrophils) were lowered, and lymphocyte and antibody production were promoted to defend against invading pathogens. Essentially, the less energy expended by chickens and pigs on the non-specific immune system, the more energy that is available for growth and production. Conclusively, phytochemicals did modulate NFκB and/or MAPKs signaling pathways in chickens and pigs, but the detailed mechanisms, including expression of upstream molecules involving NFκB and MAPKs signaling as well as the relationship between antioxidant and anti-inflammation, are still under investigation.
ACKNOWLEDGMENTS
The authors would like to express the sincere gratitude to the Ministry of Science and Technology (MOST 106-2313-B-005-048) and the iEGG and Animal Biotechnology Center for financially supporting the study to Prof. Tzu-Tai Lee.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.