The effects of low-protein diets and protease supplementation on broiler chickens in a hot and humid tropical environment
Article information

Abstract
Objective
This experiment was conducted to investigate the effects of dietary crude protein (CP) level and exogenous protease supplementation on growth performance, serum metabolites, carcass traits, small intestinal morphology and endogenous protease activity in broiler chickens reared under a tropical climate.
Methods
A total of 480 day-old male broiler chicks were randomly assigned to eight dietary treatments in a 4×2 factorial arrangement. The main effects were CP level (21.0%, 19.7%, 18.5%, or 17.2% from 1 to 21 days and 19.0%, 17.9%, 16.7%, or 15.6% from 22 to 35 days) and protease enzyme supplementation (0 ppm or 500 ppm). All experimental diets were fortified with synthetic feed-grade lysine, methionine, threonine and tryptophan to provide the minimum amino acid recommended levels for Cobb 500.
Results
Reducing dietary CP linearly reduced (p<0.05) growth performance, serum albumin, total protein, and carcass traits and increased (p<0.05) serum triglycerides and abdominal fat. There was no consistent effect of reducing dietary CP on morphological parameters of the intestine and on the pancreatic and intestinal endogenous protease activity (p>0.05). Protease supplementation improved (p<0.05) feed conversion ratio, body weight gain, carcass yield and intestinal absorptive surface area.
Conclusion
Protease supplementation, as measured by growth performance, intestinal morphology and carcass yield, may alleviate the detrimental effects of low protein diets in broiler chickens.
INTRODUCTION
Cost minimization and environmental concern are consistently driving poultry nutritionists to improve the efficiency of feed utilization by formulating digestible, practical diets to avoid overfeeding with nutrients. Over the past few decades, numerous studies have been published on the use of synthetic amino acids (AAs) in low protein (LP) diets [1,2,3]. The supplementation of AAs in these studies varied widely depending on the type of supplemented AA, such as first and second limiting AA, feed-grade AAs, all essential amino acids (EAAs) or both EAAs and non-essential amino acids (NEAAs). However, usage of LP diets in some excessive levels still has a negative impact on growth performance, carcass traits [1], morphology of intestinal villi [4] and profitability. The scenario is more challenging when broilers are raised under high-temperature conditions, either during summer in temperate regions or in a tropical climate [5]. Although one study suggested LP diets as a means of reducing metabolic heat production associated with protein catabolism [6], others suggested the use of higher dietary crude protein (CP) mainly due to lower feed intake (FI) and protein digestibility under high-temperature conditions [7]. It is possible that the combination of these two strategies results in better performance by providing an LP diet supplemented with commercially available feed-grade AA and exogenous enzymes [8–10].
Although use of exogenous enzyme blends (i.e., carbohydrases, phytase, and protease) is well-established and widely accepted to improve digestibility of nutrient elements [8,10], use of single proteases is still under investigation for widespread application. Single exogenous protease, when added to the diet, effectively increased the apparent AA and protein digestibility in the broiler chicken’s diet, even in an LP diet [11,12]. However, some reports did not confirm the conversion of those improvements in broiler performance [13,14]. Proteases may have an ‘extra-proteinaceous’ advantage for broilers reared under high temperature conditions by improving gut health [15]. Supplemental mono-component protease reduced the population of intestinal clostridium [16] and improved intestinal villi height (VH) and VH/crypt depth (CD) ratio [17] in broiler chickens.
The health and development of the intestinal epithelium and its absorptive surface area (ASA) are reported to be diminished under heat stress conditions, leading to poorer nutrient absorption and performance in broiler chickens [18]. Thus, the current study was undertaken to explore the effect of exogenous microbial protease supplementation on growth performance, blood metabolites, carcass traits and intestinal morphology in broiler chickens that were fed LP diets and reared under a hot and humid tropical climate.
MATERIALS AND METHODS
Experimental design and diets
Four isocaloric diets with descending levels of CP namely i) 21.0% (day 1 to 21) and 19.0% (day 22 to 35), ii) 19.7% (day 1 to 21) and 17.9% (day 22 to 35), iii) 18.5% (day 1 to 21) and 16.7% (day 22 to 35), iv) 17.2% (day 1 to 21) and 15.6% (day 22 to 35) were formulated. Diets were fortified with synthetic feed-grade lysine, methionine, threonine and tryptophan to provide the minimum AA recommended levels for Cobb 500 (Table 1). Four hundred and eighty birds were randomly assigned to diets without or with microbial protease (300,000 units/kg) supplementation from day 1 to 35. Thus, the experiment had a 4×2 factorial arrangement of treatments with diets containing four levels of CP and two levels of enzyme supplementation. There were four replicate pens per dietary CP-enzyme supplementation subgroup.
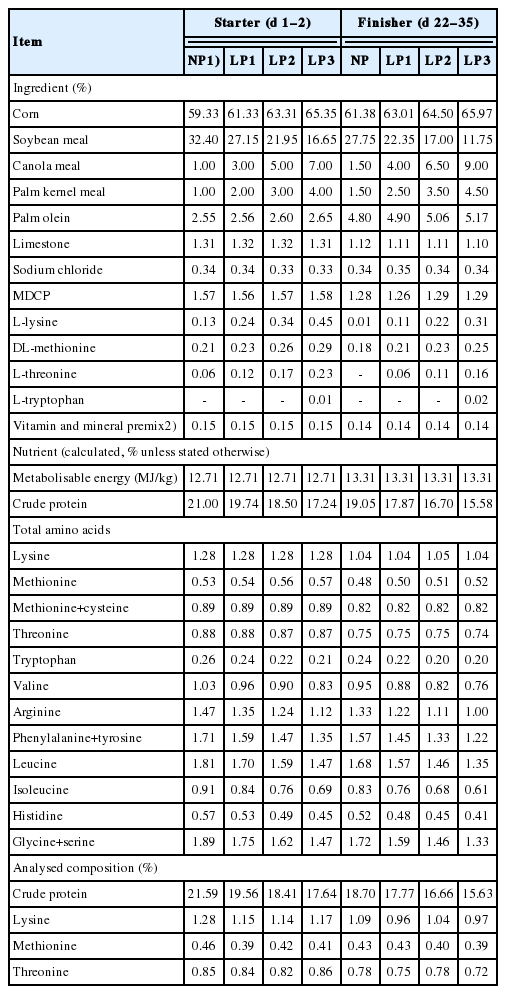
Ingredient and nutrient compositions of experimental diets
Enzyme composition
A purified microbial protease (Cibenza DP100; Novus International Inc., St. Charles, MO, USA) was used in this study. The enzyme is an alkaline serine endopeptidase protease derived from Bacillus lichenformis with a protease activity of 600,000 units/g. The manufacturer’s recommended inclusion rate is 300 units/g of feed. The protease activity was determined using the method described by Jin et al [19]. The protease activity in protease-added diets for NP, LP1, LP2, and LP3 diets were 324, 294, 325, and 327 units per gram of starter feed and 305, 325, 286, and 321 units per gram of finisher feed, respectively.
Birds and management
All experimental procedures were conducted in accordance with Universiti Putra Malaysia Research Policy on Animal Ethics and Welfare. A total of 480 day-old male Cobb 500 broiler chicks (46.56 g±0.01) were randomly distributed to 32 floor pens (n = 15 chicks/pen) with wood shavings that serve as deep litter material in a conventional open-sided poultry house and under the hot and humid tropical conditions. The temperature ranged from 23°C to 36°C, and the relative humidity was between 75% and 90%. Respective diets (mash feed) and water were provided ad libitum from day (d) 1 to 35.
Data collection and sampling
Birds and feed were weighed by pen group at the initial of the experiment (d 1) and the remaining of the feed and body weight were recorded on d 1, 21, and 35. Feed supple and left over of the feed consumption were weighed during the same period. The FI data was adjusted for mortalities and feed conversion ratio (FCR) was calculated as ratio of feed consumption to weight gain. Mortality was recorded as it occurred. On d 35, two birds per replicate were randomly selected, individually weighed and sacrificed for collection of blood, intestinal tissue and digesta. Blood samples were centrifuged at 3,000 g at 4°C for 15 min to obtain the serum. The haemolysis-free serum was transferred immediately and frozen at −80°C until further biochemical analyses. For measurement of intestinal morphology, approximately 5 cm of the middle portion of the duodenum (apex section), jejunum (between the entry of bile ducts and Meckel’s diverticulum) and ileum (between Meckel’s diverticulum and caecal junction) were cut, gently flushed with 0.9% saline solution, and placed in 10% neutral-buffered formalin. Additionally, intestinal digesta contents from the distal end of duodenum to ileo-caecal junction and pancreas samples were collected for protease activity measurement. The pancreas and intestinal content samples were collected from each bird within 5 min after death, placed in a 5-mL screw-capped tube and immediately frozen in liquid nitrogen and then stored at −80°C until further analysis.
Carcass traits
On d 36, two birds, based on the average body weight per pen, were randomly selected (eight birds/dietary treatment), leg-banded, and weighed individually. Birds were slaughtered by the Halal slaughtering process. When the birds were completely immobilized, carcasses were scalded at 55°C to 60°C for 45 s and defeathered in a rotary plucker. The carcass weight was recorded with abdominal fat, lungs and kidneys but without giblets, feet or the head. Non-deboned breast meat with the attached skin (both pectoralis major and minor) and abdominal fat were removed and weighed. The abdominal fat pad was considered the tissue surrounding the gizzard and intestines, extending within the scheme, neighbouring the cloaca and bursa of fabricius, and adjoining the abdominal muscle.
Gut morphology
The intestinal segments (2 to 4 mm) were dehydrated for 16 h in an automatic tissue processor (Leica ASP 3000; Leica Biosystems GmbH, Nussloch, Germany) before being embedded in paraffin (Leica EG 1160; Leica Biosystems GmbH, Germany). Duplicate tissue sections with 4-μm thickness were produced using a rotary microtome machine (Leica RM 2155; Leica Biosystems GmbH, Germany). The sections were placed on a glass slide and heated at 57°C until dried. Upon drying, the slides were stained with haematoxylin-eosin and covered with a cover-slip. Histological indices were analysed under a computer-aided light microscope using image analysis (Leica Camera AG, Solms, Germany). The VH (from the crypt mouth to the villus tip), CD (from the base up to the region of transition between the crypt and villi) and villus width (VW) (half of the villus length) were measured. Accordingly, the apparent ASA (VH×VW) and the VH/CD ratio were calculated [4].
Endogenous protease activity
Pancreas tissues were processed by adding five times their weight in ice-cold 0.2 M Tris buffer (pH 8.0) with 0.05 M NaCl and then homogenized using Ultra-Turrax homogenizer (IKA Works Inc., Wilmington, NC, USA). The homogenates were centrifuged at 3,000×g for 15 min at 4°C, and the supernatant was used for protease activity measurement as described by Jin et al [19]. Digesta samples were diluted 10 times (w/v) using ice-cold phosphate-buffered saline (pH 7.0) and then homogenized and sonicated for one min with three cycles at 30 s intervals. The samples were later centrifuged at 18,000×g for 20 min at 4°C, and the supernatants were used for the protease enzyme activity assay [19]. All procedures were handled on ice to prevent possible enzyme degradation. The protease activity unit was defined as milligrams of azocasein degraded during 2 h of incubation at 38°C per mg of intestinal digesta protein (U/mg protein) or pancreas. The intestinal digesta protein concentrations were determined using colorimetric reaction of the bicinchoninic acid protein assay kit (Sigma-Aldrich, St. louis, MO, USA).
Biochemical analysis
Feed samples were collected and finely ground using a coffee grinder (Panasonic, Shah Alam, Selangor, Malaysia). The dry matter (DM) and CP were determined following the AOAC methods [20], respectively. The AA concentrations were determined using pre-column derivatization with ACCQ reagent (6-aminoquinolyl-N-hydroxysuc-cinimidyl carbamate, Waters, Milford, MA, USA) and high-performance liquid chromatography [5]. Cysteine and methionine were analysed as cysteic acid and methionine sulfone by oxidation with performic acid for 16 h at 0°C. Serum concentrations of total protein (TP), albumin (ALB), uric acid (UA), and triglycerides (TG) were analysed by an automated chemistry analyser (Hitachi 902 Automatic Analyser; Hitachi, Tokyo, Japan) using commercial test kits (Roche Diagnostics, Basel, Switzerland).
Statistical analysis
The effects of CP level, enzyme supplementation and their interactions with growth performance, serum biochemical profile, carcass traits, intestinal morphology, and pancreatic and intestinal protease activity were analysed using the general linear models procedure in SAS (SAS Institute Inc., Cary, NC, USA). Duncan’s multiple range test was used to compare between means. When interactions were significant, comparisons were made within each experimental variable. The effect of decreasing protein level was portioned into linear and quadratic components using orthogonal polynomial contrasts. Mortality data were subjected to a chi-square test. The significance is considered when p<0.05.
RESULTS
Growth performance
The effects of CP level and protease supplementation on FI, weight gain (WG), and FCR are presented in Table 2. There were no significant (p>0.05) CP×protease interactions for FI, WG, or FCR except FI for d 29 to 35. Except during the first week of life, FI was not affected by CP level during week 2 (p = 0.07) and 3 (p = 0.523). The FI was reduced linearly during d 1 to 7 (p<0.001) and d 22 to 28 (p<0.001) and d 29 to 35 (p = 0.020). The weekly data also showed that WG decreased linearly and FCR increased linearly by reduction of dietary CP. However, the results of WG and FCR were not significant for d 29 to 35. The FI and WG decreased linearly (p<0.001; p<0.001) and FCR increased linearly (p<0.001) with the reduction in dietary CP levels during d 1 to 35. The effect of protease supplementation was negligible on WG and FCR during the first 2 weeks of life. But thereafter the protease supplementation improved WG and FCR. Irrespective of dietary protein level, protease supplementation improved WG and FCR during d 1 to 21 (p = 0.013; p = 0.002) and d 1 to 35 (p<0.001; p = 0.001). The FI was not affected (p>0.05) by supplemental protease. Neither CP level nor enzyme supplementation had a significant effect on mortality rate (Table 2).
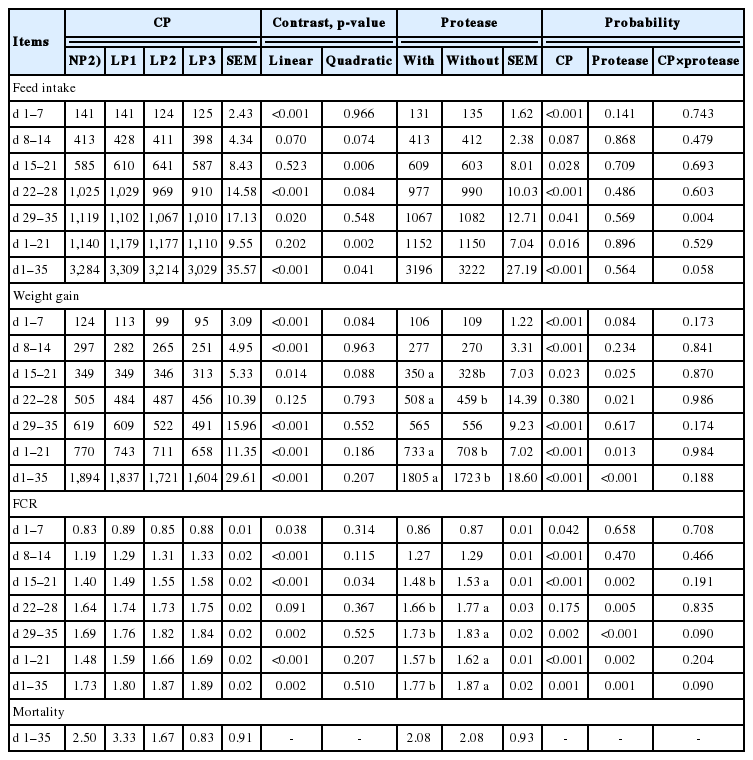
Serum biochemical profile
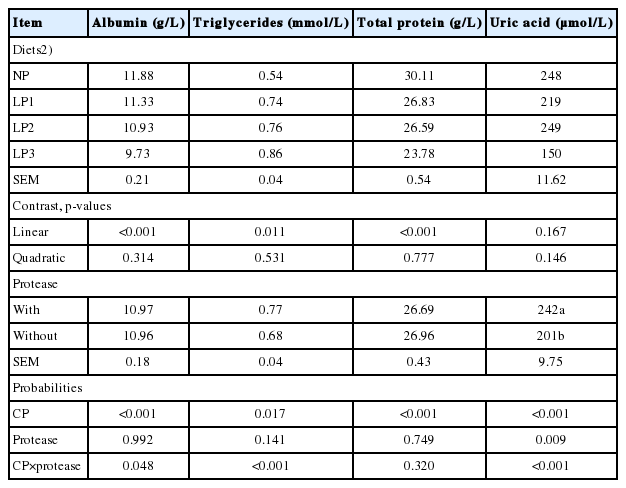
The effect of dietary CP level and protease supplementation on serum biochemicals is shown in Table 3. CP×protease interactions were significant for serum levels of ALB, TG, and UA but not for serum TP. Irrespective of protease supplementation, reducing the CP level linearly decreased (p<0.001) serum TP. Protease supplementation did not affect serum TP (p = 0.749). Serum ALB was linearly decreased by the dietary CP level reduction in both diets with and without protease (p< 0.001, p = 0.042, respectively) but was otherwise noted for TG and UA (Table 4). The level of TG increased linearly (p<0.001) with the reduction of dietary CP in the protease supplemented group. Within the non-protease-supplemented group, the level of UA decreased linearly (p<0.001) and quadratically (p<0.07).

Carcass traits
The CP level×protease interactions for carcass traits were not significant (Table 5). Reducing dietary CP linearly decreased the dressed yield (p<0.001) and breast meat (p = 0.049) and conversely increased the abdominal fat (p<0.001), regardless of protease supplementation. Protease supplementation increased carcass yield (p = 0.042), irrespective of dietary CP level. However, percentages of breast meat and abdominal fat pad were unaffected by protease supplementation.

Gut morphology
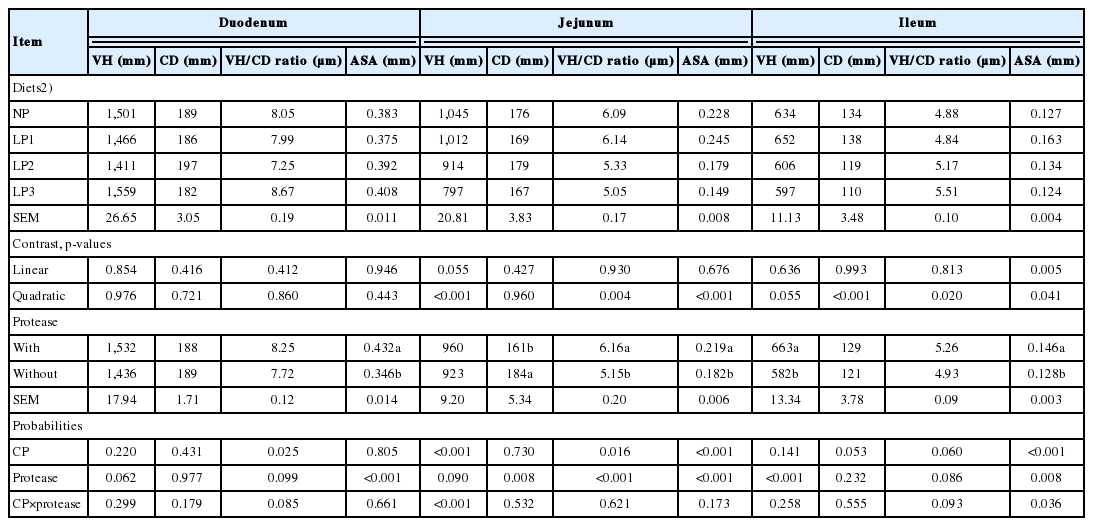
The effects of dietary CP level and protease supplementation on the VH, CD, VH/CD ratio and ASA in the duodenum, jejunum and ileum are displayed in Table 6. No significant CP×protease interactions were noted for duodenal VH, CD, VH/CD, and ASA, jejunal CD, VH/CD ratio and ASA and ileal VH, CD and VH/CD. However, significant CP level× protease interactions were found for jejunal VH and ileal ASA (p = 0.036). Irrespective of CP level, protease supplementation significantly increased duodenal ASA (p<0.001), jejunal VH/CD ratio (p<0.001) and ASA (p<0.001) and ileal VH (p<0.001). However, protease supplementation reduced jejunal CD (p = 0.008). Regardless of protease supplementation, jejunal VH (p<0.001), VH/CD (p = 0.004) and ASA (p<0.001) and ileal CD (p<0.001) decreased quadratically. However, ileal VH/CD increased quadratically (p = 0.020) with the reduction in dietary CP. In contrast, reducing dietary CP had no effect on duodenum VH/CD and ASA (p>0.05). Jejunal VH decreased linearly (p<0.001) and quadratically (p<0.001) with protease in LP diets, but only a quadratic decrease was observed when protease was supplemented. Ileal ASA increased linearly (p = 0.001) when protease was added to LP diets, whereas a quadratic decrease (p = 0.004) was observed among those without supplemental protease (Table 7).

Enzyme activity
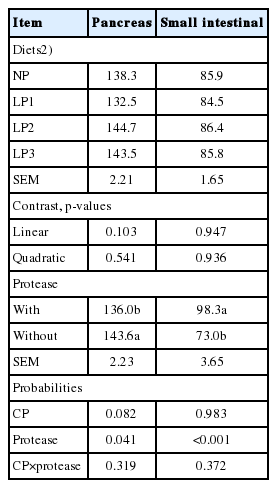
No significant CP level×protease interactions were observed in pancreatic or intestinal protease activity (Table 8). Protease supplementation increased (p<0.001) intestinal protease activity and decreased (p = 0.041) pancreatic protease activity. Neither pancreatic nor intestinal protease activity was affected by dietary CP level.
DISCUSSION
Hot and relatively humid environmental constraint can generate a state of physiological and behavioural stress that might affect the nutrient requirement in broiler chickens which evoke a negative effect on their productive performance, immunity and survivability [7]. Beneficial use of LP diets with supplemental EAA in a hot-weather has been reported [5,6]. In the present study, despite meeting the requirement for lysine, methionine+cysteine, tryptophan and threonine in LP diets, WG and FCR were depressed throughout the experimental period, regardless of protease supplementation. Reduction of growth performance in broilers fed LP diets and supplemented with EAA has been reported [5] irrespective of their environmental condition, and this reduction could be associated with lower levels of specific NEAA such as glycine and serine or glutamic acid, or with anon-specific need for nitrogen [6]. Earlier work on AA-supplemented LP diets in a hot environment [5] suggested that reduced FI could be a possible explanation for the inferior WG. The reduced FI was noted as a quadratic effect in this study, where only higher levels of CP reduction (LP2 and LP3) reduced FI markedly. This may possibly be due to one or several AA that are regulating FI other than CP concentrations per se [21]. Although protease supplementation improved WG and FCR, it failed to alter the linear reduction of WG and the increment of FCR incurred by reducing CP. This could be partly due to the lack of protease effect on FI. The protease used in this study was previously shown to enhance proteins, AAs and energy digestibility in broilers fed different protein levels [12]. Therefore, the observed improvements in nutrient digestibility and nutrient retention [15,17,22] in birds fed diets containing exogenous protease may have beneficial effects on body weight. This finding is in accordance with those reported by Angel et al [11] and Mahmood et al [10], where supplementation of mono-component protease improved growth performance in broilers.
The data showed that descending levels of dietary CP reduced the concentrations of ALB and TP, irrespective of protease supplementation. Blood plasma TP and ALB are the main transport proteins in avian species, and they reflect the avian nutritional condition [23]. The reduction in serum TP and ALB could be attributed to deficiency in AA intake by the birds, as measured by the lower FI [5]. The linear reduction of FI (day 1 to 35) in our study may have accounted for the decreased UA level with the reduction in dietary CP level. The UA is a by-product of protein catabolism and turnover in the body. Thus, the decrease in UA could be associated with a deficiency of ingested AA. Furthermore, UA reduction may also be related to glycine deficiency, the substrate of UA synthesis in the liver [2,3]. It is interesting to note that the linear reduction in UA with decreasing dietary CP level was only noted among those without protease supplementation. It appears that protease supplementation may have released glycine in the digestive system and therefore compensated the shortage of glycine in LP diets [12]. On the other hand, serum TG was not affected by reduced protein diets containing no protease. However, decreasing CP with supplemental protease increased serum TG. This finding is in contrast to the findings of Abudabos [24]. The author fed broiler chickens with a commercial enzyme cocktail (protease and carbohydrase) and observed that enzyme supplementation has no effect on serum TG. This discrepancy may be due to the different types of enzyme used.
As expected, carcass and breast meat yield decreased, and the rate of abdominal fat deposition was significantly higher in birds that were fed LP diets in this study. Most of the previous studies reported the same phenomenon in broilers fed with LP diets [3,24]. This is due to a higher calorie:protein ratio in LP diets. It appears that the excess available energy beyond that required for protein deposition is diverted to abdominal fat deposition. Interestingly, although protease supplementation did not change the rate of fat deposition, it improved the carcass yield. There are conflicting reports on the effect of protease supplementation on carcass yield. While Rehman et al [22] noticed no significant change in carcass yield or abdominal fat, Abudabos [24] reported an increase in carcass and breast meat yield. The improvement in carcass yield associated with protease supplementation could be due to enhanced utilization and deposition of protein [23].
The present results concur with earlier work that reducing dietary CP was detrimental to gastrointestinal tract morphometry [4]. The decrease in the intestinal VH, VH/CD, and ASA is associated with poor nutrient absorption and consequently depressed growth performance in broiler chickens [17]. The adverse effect of LP diets on gut morphometry could be attributed to a decrease in the concentrations of NEAA such as glycine, glutamine and proline, which actively support the epithelial layer and represent a major fraction of intestinal masons and digestive secretions [25]. In contrast, protease supplementation significantly increased duodenal and jejunal ASA and jejunal VH/CD ratios and ileal VH. The present findings are in agreement with Cowieson et al [15], Ding et al [9] and Xu et al [17]. These results indicated that more nutrients were digested by supplemental protease [9,11], and most likely, the extra energy absorbed was used to improve the intestinal morphology.
It was noted that the reduction in dietary CP level did not alter pancreatic or intestinal protease activity in the present study. Ding et al [9], however, reported otherwise. On a cautionary note, Ding et al [9] reduced CP (21%, 20%, and 19% during starter and 19%, 18%, and 17% during grower phase) and supplemented with AA (lysine, threonine, and methionine). However, the calculated and analysed AA were lowered in LP diets. Thus, the discrepancy between pancreatic and intestinal protease activities may be due to insufficient supplementation of AA. The current results confirmed those of Mahagna et al [13], which showed that enzyme supplementation (protease and amylase at higher dosage, 1,000 ppm) suppressed endogenous pancreatic protease activity. Exogenous proteases may augment endogenous peptidases by increasing protein digestibility and hydrolysing proteinaceous anti-nutritional factors. In contrast, Ding et al [9] fed broiler chickens with three levels of protease (0, 150, and 300 ppm) and observed that protease concentration has no effect on either pancreatic or small intestinal protease activity at d 21. However, protease (at 300 ppm) significantly increased pancreatic protease activity at d 42. These inconsistencies could be attributed to variations in the age of birds, the dosage, or the type of protease used [26].
CONCLUSION
The present findings demonstrated that protease supplementation at the rate used in this study may offset the detrimental effects caused by reducing dietary CP on growth performance, intestinal morphology and carcass yield in broiler chickens raised under a hot and humid tropical condition. The magnitude of compensation may depend on the type and dosage of protease supplemented.
ACKNOWLEDGMENTS
This study was supported by the Malaysian Ministry of Higher Education under the Long-term Research Grant Scheme.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.