Choline Essentiality and Its Requirement in Diets for Juvenile Parrot Fish (Oplegnathus fasciatus)
Article information
Abstract
A 12-wk feeding trial was conducted to evaluate the essentiality of choline supplementation in diets for parrot fish. Five isonitrogenous and isocaloric diets were supplemented with 0 (as control), 500, 1,000, and 2,000 mg choline per kg diet, and a positive control diet without choline contained 0.3% of 2-amino-2-methyl-1-propanol as choline biosynthesis inhibitor (designated as Con, C500, C1000, C2000 and Con+, respectively). Triplicate groups of fish (body weight, 8.8±0.01 g) were fed one of the experimental diets at a rate of 4% body weight twice daily. The fish fed Con+ diet revealed significantly lower growth performance and feed utilization efficiency than other fish groups. Supplementation of choline to the basal diet did not significantly influence fish growth. The highest liver lipid content was observed in fish fed the Con+ diet and inversely correlated with liver choline concentration although the differences were not significant. Also, significantly higher liver linoleic, eicosapentaenoic and docosahexaenoic acid contents were found in fish fed the Con+ diet. Innate immune parameters including respiratory burst and myeloperoxidase activities were not significantly affected by dietary choline levels. The findings in this study conclude that choline concentration of approximately 230 mg kg−1 diet meets the requirement of parrot fish.
INTRODUCTION
Choline is an essential component of cell structure and plays important roles in cell maintenance and certain metabolic functions (McDowell, 2000). It mostly exists in the forms of phospholipids phosphatidylcholine (lecithin), lysophosphatidylcholine, choline plasmalogens and sphingomyelin, and is widely distributed in all plant and animal cells (Zeisel, 1990). It is also an essential component of acetylcholine, the most common neurotransmitter in the nervous system, which is involved in transmission of nerve impulses across synapses (Wauben and Wainwright, 1999). The prominent structural feature of choline is its biologically active methyl groups, through which plays a vital role as a methyl donor in synthesis of various methylated metabolites (Combs, 1992). Furthermore, choline is considered as a lipotropic factor preventing abnormal accumulation of lipid and development of fatty livers (Halver, 2002).
From another point of view, choline is a vitamine-like nutrient and is regarded as an essential dietary component for most young vertebrates (NRC, 2011) including fish; nevertheless it can be synthesized in liver from other methyl donors such as methionine. Most animals can synthesize choline through the methylation of phosphatidyl-ethanolamine mediated by phosphatidylethanolamine-N-methyltransferase (PEMT) (McDowell, 2000). The PEMT catalyzes the methylation of phosphatidylethanolamine to form phosphatidylcholine (PC) and represents one of the two major pathways for PC biosynthesis. However, it is believed that the rate of de novo choline synthesis is not sufficient or rapid enough to meet the metabolic requirement of animal including juvenile fishes when the diet is deficient in methyl group donors (Wilson and Poe, 1988).
The signs of choline deficiency including growth retardation, poor survival, poor feed efficiency (FE) and increased liver lipid concentration have been reported in carp (Cyprinus carpio) (Ogino et al., 1970), lake trout (Salvelinus namaycush) (Ketola, 1976), rainbow trout (Oncorhynchus mykiss) (Rumsey, 1991), hybrid striped bass (More saxatilis×Morone chryso) (Griffin et al., 1994), red drum (Sciaenops ocellatus) (Craig and Gatlin, 1996), yellow perch (Perca flavescens) (Twibell and Brown, 2000), channel catfish (Ictalurus punctatus) (Zhang and Wilson, 1999), hybrid tilapia (Oreochromis niloticus×O. aureus) (Shiau and Lo, 2000) and cobia (Rachycentron canadum) (Mai et al., 2009). Dietary choline requirement has been estimated for several fish species and the value has been reported to be at the range of 400 to 1,000 mg/kg diet.
Parrot fish is a carnivorous species and is regarded as an emerging aquaculture species due to its high economic value, excellent flesh quality and diseases resistance. However, there are just a few available studies on nutrient requirement of the species (Wang et al., 2003; Nam et al., 2005; Shan et al., 2008; Kim et al., 2009; Lim and Lee, 2009; Bueno Galaz et al., 2010; Lim et al., 2013). Therefore, the present study was conducted to evaluate the dietary choline requirement and its essentiality for parrot fish.
MATERIALS AND METHODS
Experimental diets
Formulation of the experimental diets is provided in Table 1. A basal experimental diet was formulated and used as a control and three other diets were prepared by supplementing 500, 1,000, or 2,000 mg/kg choline (designated as Con, C500, C1000, and C2000, respectively). Another control diet without choline supplementation was prepared to contain 0.3% of 2-amino-2-methyl-1-propanol (AMP) as a choline biosynthesis inhibitor and regarded as positive control (Con+). The analyzed choline concentrations of the experimental diets were 230 (Con), 230 (Con+), 663 (C500), 1,000 (C1000), and 1,554 (C2000) mg/kg diet respectively. White fish meal (Suhyup Feed Co. Ltd., Uiryeong, Korea) and casein were used as dietary protein sources. To remove choline from fish meal, it was extracted two times with 70% aqueous ethanol solution (ethanol/water, 7/3, v/v) for 48 h, and then dried using an electric fan at room temperature. The dietary ingredients were thoroughly mixed with squid liver oil and 30% distilled water in a mixer (NVM-14-2P, Busan, Korea). The wet dough was pelleted by a chopper machine (SMC-12, Kuposlice, Busan, Korea) at 4 mm of diameter. The diets were then freeze-dried at −40°C for 24 h, crushed into desirable particle sizes (1 to 3.0 mm) and stored at −45°C until used.

Formulation and proximate composition of the experimental diets (% dry matter)
Fish and feeding trial
Juvenile parrot fish were transported from a private hatchery (Chang-Hae Fisheries Co., Jeju-Island, Korea) to the Marine and Environmental Research Institute (Jeju National University, Jeju, Korea). The fish were fed a commercial diet for one month to become acclimated to the experimental facilities and conditions. At the end of the acclimation period, the fish (body weight, 8.8±0.01 g) were randomly assigned to fifteen 150-L polyvinyl circular tanks at a density of 25 fish per tank. The experiment was done in triplicates and the fish were fed at a rate of 4% body weight twice daily (8:00 and 18:00 h) for 12 wk. A flow-through system was used and the tanks were supplied with sand filtered seawater at a rate of 3 L/min and aeration to maintain enough dissolved oxygen. The photoperiod was scheduled by 11:13 h (light/dark) using fluorescent light. Water temperature ranged from 19°C to 25°C according to the seasonal changes. Salinity of the water was 32‰ to 34‰, dissolved oxygen ranged from 7.80 to 8.05 mg/L, and the pH was maintained at 8.02±0.01. The growth of fish was measured every two wk and feeding rate was adjusted accordingly. The fishes were fasted for 24 h prior to weighing or blood sampling. The tanks were routinely cleaned using sponge to prevent the growth of microflora. Experimental protocols followed the guidelines approved by the Animal Care and Use Committee of Jeju National University.
Sample collection and analyses
At the end of the feeding trial, all the fish in each tank were weighed and counted for calculation of growth performance and survival. Three fish per tank were sampled for measurement of liver lipid and choline concentrations and fatty acid composition. Also, another set of three fish per tank (9 fish per treatment) were randomly selected and anaesthetized with 2-phenoxyethanol (100 mg/L), and blood samples were taken from the caudal vein using non heparinized syringes. The samples were used for determination of respiratory burst and myeloperoxidase (MPO) activities. All the analyses were performed in Fish Feed and Nutrition Laboratory of Jeju National University (Jeju, Korea).
The liver lipid concentration was analyzed using the method of Folch et al. (1957). Choline concentration in the experimental diets and liver samples were measured using the choline/acetylcholine assay kit (Abcam, UK).
The liver lipid was extracted according to Folch et al. (1957). Fatty acid methyl esters (FAMEs) of liver lipids were prepared using acid-catalysed transesterification method (Metcalfe et al., 1961). The FAMEs were analyzed by gas chromatography (GC; HP 6890, Agilent, Wilmington, DE, USA) using an HP-88 column (100 m×250 μm×0.25 μm; Agilent, USA). The GC conditions were as follows: Injector and detector temperature was 260°C; carrier gas, H2; and split ratio, 1/100. The programmed temperature was as follows: 140°C for 5 min; increase of 4°C/min to 230°C; 230°C for 10 min; increase of 3°C/min to 240°C; and 240°C for 10 min. Peaks were identified by comparison of retention times with those of the corresponding standards of FAMEmix-37 (Supelco, Bellefonte, PA, USA). Fatty acids concentrations were expressed as percentage (w/w) of total FAMEs.
Monitoring of non-specific immune responses
Oxidative radical production by phagocytes during respiratory burst was measured through nitro-blue-tetrazolium (NBT) assay described by Anderson and Siwicki (1995). Briefly, blood and NBT (0.2%) (Sigma, St. Louis, MO, USA) were mixed in equal proportion (1:1) and incubated for 30 min at room temperature. Then 50 μL was taken out and dispensed into glass tubes. One ml of dimethylformamide (Sigma, USA) was added and centrifuged at 2,000×g for 5 min. Finally, the optical density of supernatant was measured at 540 nm using a spectrophotometer (Genesys 10UV, Rochester, NY, USA). Dimethylformamide was used as the blank.
The MPO activity was measured according to Quade and Roth (1997). Briefly, 20 μL of serum was diluted with Hanks balanced salt solution without Ca2+ or Mg2+ (Sigma-Aldrich, USA) in 96-well plates. Then, 35 μL of 3,3′,5,5′-tetramethylbenzidine hydrochloride (20 mM) (Sigma-Aldrich, USA) and H2O2 (5 mM) were added. The color change reaction was stopped after 2 min by adding 35 μL of 4 M sulfuric acid. Finally, the optical density was read at 450 nm in a microplate reader.
Statistical analysis
All experimental diets were assigned by a completely randomized design. Data were subjected to one-way analysis of variance (ANOVA) in SPSS version 11.0 (SPSS Inc., Chicago, IL, USA). The significant differences between group means were compared using Tukey’s honestly significant difference at the 5% level of significance (p<0.05). Data are presented as mean±standard deviation. Percentage data were arcsine-transformed before the ANOVA analysis.
RESULTS
The results of fish growth performance and feed utilization are shown in Table 2. The results showed a significant decrease of growth performance in fish fed the Con+ diet in comparison to the other groups. Also, a significant reduction in FE was observed in the group offered the Con+ diet. No significant effects of choline supplementation were found on growth or feed utilization although numerically higher values were recorded for fish fed C1000 and C2000 diets compared to those fed the Con diet. Feed intake did not significantly differ among the experimental groups. The survival rate varied from 96% to 98% without any significant differences.

Growth performance of parrot fish (initial body weight 8.80±0.01 g) fed the five experimental diets for 12 wk
Innate immune parameters including NBT and MPO activities did not significantly differ among dietary treatments (Table 3).

Non-specific immune response of parrot fish fed the five experimental diets for 12 wk
The highest liver lipid concentration was observed in fish fed Con+ diet, but no significant differences were detected (Figure 1). The liver choline concentration showed a positive correlation with dietary choline level and inversely correlated with liver lipid content. Liver fatty acid composition of fish fed the diets is shown in Table 4. Fish fed the Con+ diet exhibited significantly higher accumulation of linoleic acid, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) in the liver compared to the other fish groups. An opposed tendency was observed for palmitic acid, where significantly lower value was obtained in fish fed the Con+ diet than those fed C1000 diet.
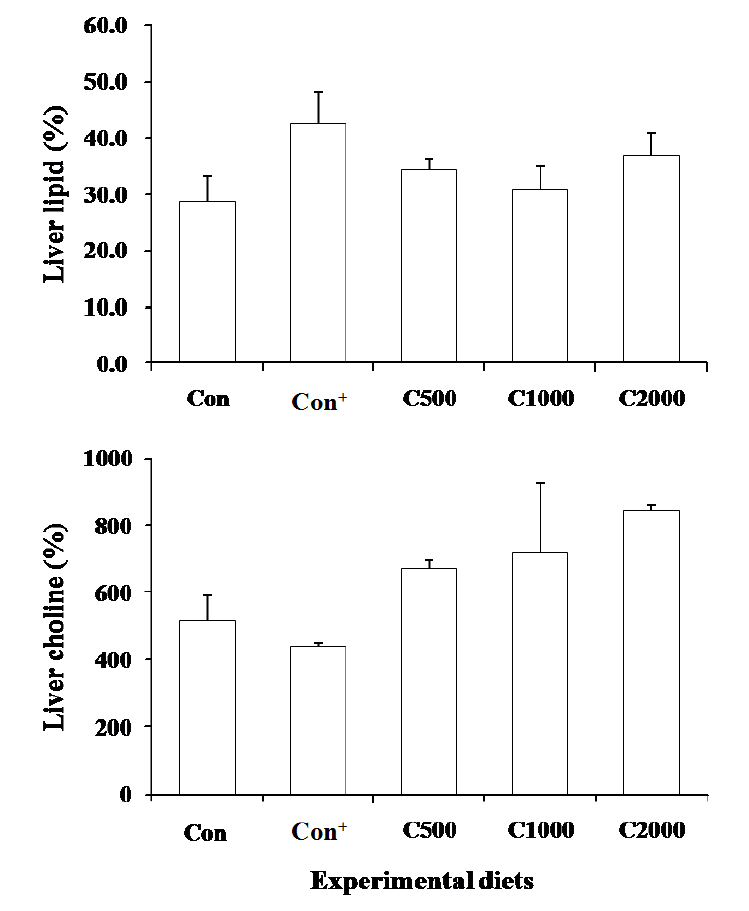
Liver lipid and choline concentration of juvenile parrot fish fed the experimental diets for 12 wk.

Liver fatty acids composition (% total fatty acids) of parrot fish fed the five experimental diets for 12 wk
DISCUSSION
In this study a reduced growth performance was obtained when AMP was included in the diet. The AMP is one of the choline antagonists which have been demonstrated to slow down the choline biosynthesis through the inhibition of S-adenosylmethionine-dependent methylation and results in choline deficiency symptoms. Rumsey (1991) reported that it can effectively block the choline biosynthesis in rainbow trout. The significant reduction in weight gain and FE of fish by AMP supplementation in this study indicated that it can effectively inhibit de novo choline synthesis in parrot fish.
Methionine, as one of the sulfur amino acids, is a well-known nutritional factor that influences choline requirement. Choline is synthesized by the sequential methylation of phosphatidylethanolamine in several animal species including fish (NRC, 1993). In this regard, methionine is involved in choline synthesis by enhancing the availability of S-adenosylmethionine which serves as methyl donor (Bender, 1992; NRC, 1993). In the present study, the Con diet contained 1.84% methionine which is probably higher than the requirement level for parrot fish. Our results showed that the fish fed Con diet had significantly higher growth than those fed AMP containing diet, and the values were comparable to those of fish fed the choline supplemented diets. This may indicate the de novo synthesis of choline in parrot fish. The strong endogenous choline synthesis has also been earlier observed in channel catfish (Zhang and Wilson, 1999), while it has been shown that a dietary source of choline is required for rainbow trout (Rumsey, 1991), hybrid striped bass (Griffin et al., 1994), yellow perch (Twibell and Brown, 2000), hybrid tilapia (Shiau and Lo, 2000) and cobia (Mai et al., 2009) when dietary sulfur amino acids concentration are not provided in excess level.
The results of previous studies on hybrid striped bass (Griffin et al., 1994), yellow perch (Twibell and Brown, 2000), hybrid tilapia (Shiau and Lo, 2000) and cobia (Mai et al., 2009) showed that the optimal choline requirement level ranges from 500 to 1,000 mg/kg diet when choline chloride was used as choline source. In the present study the basal diet contained around 230 mg/kg choline and additional supplementation levels did not result in improvement of growth performance, indicating that the basal level of choline meets the requirements of parrot fish. The dietary content of precursors such as methionine or betaine, age/size, growth rate and species specific differences have been regarded as effective factors on response to dietary supplementation of choline (Wilson and Poe, 1988; Rumsey, 1991; Griffin et al., 1994).
Liver lipid content has been used as an indicator for evaluation of choline status in animals (McDowell, 2000; Halver, 2002). Choline as a lipotropic factor involves in metabolism and mobilization of liver lipids and prevents the abnormal accumulation of fat and development of fatty liver. It is involved in the synthesis of phospholipids such as PC, as an integral part of lipoproteins which are important in the extracellular transport of lipids in the blood and lymph (Tocher, 1995; 2003). Apparently, PC is the predominant phospholipid class in fish lipoproteins (Nelson and Shore, 1974; Chapman, 1980). The increment of liver lipid in the presence of AMP in this study may be attributed to the reduction of PC as a result of impaired lipoprotein secretion and subsequent accumulation of triglycerides. Choline supplementation to the basal diet did not influence the liver lipid content which is in agreement with earlier studies on rainbow trout and yellow perch when they were fed the diets with different levels of choline (Rumsey, 1991; Twibell and Brown, 2000). However, liver lipid content of channel catfish and hybrid striped bass significantly decreased at increased dietary choline levels (Griffin et al., 1994; Zhang and Wilson, 1999). In contrast, studies on red drum and hybrid tilapia have shown increased levels of liver lipid in response to dietary choline supplementation (Craig and Gatlin, 1996; Shiau and Lo, 2000).
In this study liver choline concentration of parrot fish was increased in a dose dependent manner. Similar results were observed in channel catfish (Zhang and Wilson, 1999) and cobia (Mai et al., 2009). Apparently, based on liver choline levels, the dietary choline requirement of parrot fish is much higher than the value based on weight gain. Mai et al. (2009) declared that this may be due to higher choline requirement for physiological functions than for growth.
Considering the fact that fatty acids are structural components of phospholipid and neutral lipid molecules which undergo altered metabolism during choline deficiency, we examined the fatty acid composition of fish liver in order to gain further insight as to the lipotropic action of choline. Interestingly, our results showed the higher accumulation of linoleic acid, EPA and DHA in the liver when the fish were provided with AMP containing diet. There is no earlier report on the effect of dietary choline on fish liver fatty acid composition. Eric et al. (1965) could not find any differences in fatty acid composition of liver between choline-deprived and choline fed rats. Watkins et al. (2003) used quantitative lipid metabolite profiling to characterize lipid metabolism in PEMT-deficient and wild mice fed diets containing varying concentrations of choline. Their results showed higher accumulation of free fatty acids in liver of mice fed the choline deficient diet compared to those fed choline supplemented diet. Also, they found that PEMT activity affects the flux of lipid between liver and plasma and the delivery of essential fatty acids to blood and peripheral tissues; where PEMT-deficient mice had substantially diminished concentrations of DHA and arachidonic acid in plasma. This finding clearly shows that impairment of endogenous choline synthesis results in accumulation of fatty acids in the liver which is in agreement with our results.
Dietary lipotropes including choline are involved in several metabolic pathways and play an important role in cellular metabolism via the regulatory effects on the transfer of one-carbon moieties which are used for DNA synthesis. Instant availability of nucleotide predecessors is especially important in lymphatic system. Therefore, deficiency of these cofactors can negatively affect the immune function and disease resistance (Nauss and Newberne, 1981). Courrèges et al. (2003) demonstrated that choline dietary deficiency produces an impairment of the immune response in rats. Their results showed that choline deficiency results in diminution of Th2/Th1 response which can lead to reduced host response to antigens. However, there is no available study on the effect of dietary choline level on fish immune response. Fishes mostly rely on non-specific immune system than specific immune system (Anderson, 1992). The non-specific immune system includes both cellular and humoral responses of which respiratory burst and MPO activities are related to the humoral immune response. During the respiratory burst reactive oxygen species are produced by phagocytes and the activity can be measured through the NBT assay. The MPO is an important enzyme with microbactericidal activity and is one of the potent agents in killing pathogens. In the present study, however, no significant differences were found among all the fish groups. The non-specific immune response lasts for a short duration and disappears with the time due to the lack of immunologic memory in this type of immunity (Anderson, 1992). Furthermore, each of innate immune factors may differently respond to the dietary additives (Secombes, 1996). Therefore, further complementary studies are needed to examine the effects of dietary choline on innate immunity of fish.
In conclusion, our results showed that choline concentration of 230 mg/kg diet largely meets parrotfish requirement for growth and normal lipid metabolism, and that the additional choline supplementation in diet is not necessary for the juvenile parrot fish. The fish seems to synthesize enough choline for its normal growth and metabolism. Further studies are required in the future to estimate the exact choline requirement of the species.
ACKNOWLEDGMENTS
This study was supported by the National Research Foundation of Korea (NRF) grant funded by the Korean Government (MEST) (No. 2011-0015925).