Effect of Alcohol Fermented Feed on Lactating Performance, Blood Metabolites, Milk Fatty Acid Profile and Cholesterol Content in Holstein Lactating Cows
Article information
Abstract
A feeding experiment with 40 lactating Holstein cows and 4 dietary treatments was conducted to investigate supplementation with different levels of alcohol fermented feed to the TMR on lactating performance, blood metabolites, milk fatty acid profile and cholesterol concentration of blood and milk. Forty Holstein lactating cows (106±24 d post-partum; mean±SD) were distributed into four groups and randomly assigned to one of four treatments with each containing 10 cows per treatment. The treatment supplemented with TMR (DM basis) as the control (CON), and CON mixed with alcohol-fermented feeds (AFF) at a level of 5%, 10% and 15% of the TMR as T1, T2 and T3, respectively. Dry matter intake and milk yield were not affected by supplementation of AFF. An increased 4% FCM in the milk occurred in cows fed T3 diet compared with CON, while T1 and T2 diets decreased 4% FCM in a dose dependent manner. Supplementation of AFF increased the concentration of albumin, total protein (TP), ammonia, and high density lipoprotein-cholesterol in serum compared with CON. In contrast, supplementation with AFF clearly decreased concentration of blood urea nitrogen (BUN) and total cholesterol (TC) compare with CON. AFF supplementation increased the proportion of C18:1n9 and C18:2n6 compared to CON. A decrease in the concentration of saturated fatty acid (SFA) for T1, T2 and T3 resulted in an increased unsaturated fatty acid (USFA) to SFA ratio compared to CON. Concentration of cholesterol in milk fat was reduced in proportion to the supplemental level of AFF. Feeding a diet supplemented with a moderate level AFF to lactating cows could be a way to alter the feed efficiency and fatty acid profile of milk by increasing potentially human consumer healthy fatty acid without detrimental effects on feed intake and milk production. A substantially decreased cholesterol proportion in milk induced by supplementation AFF suggests that alcohol fermented feed may improve milk cholesterol levels without any negative effects in lactating cows.
INTRODUCTION
Alcohol is one of the important intermediates formed during fermentation of forage and agriculture by-products. A wide range of alcohol concentrations can be found in grass and maize silages (Dulphy and Demarquilly, 1981; Byers et al., 1982; Rooke et al., 1988). Alcohol, introduced in the rumen by diets, is known to be only partially (about 20%) transformed to VFA by the rumen microorganisms (Durix et al., 1991) and most alcohol is known to be absorbed through the rumen wall (Burning and Yokoyama, 1988). In the rumen, alcohols also can be synthesized by fungi (Teunissen et al., 1992) and bacteria (Laukova and Marounek, 1992), and production of alcohol is an electron sink in fermentation (Teunissen et al., 1992), in agreement with the observed accumulation of alcohols with high starch intake (Allison et al., 1964). Alcohols are removed from the rumen by microbial metabolism and absorption (Jean-Blain et al., 1992).
The addition of an adequate amount of alcohol to beef diets has been known to be desirable to improve feed efficiency and meat quality. Shin et al. (1995) observed a higher alcohol concentration in steers fed alcohol fermented feed (AFF), and it was speculated that there was an amount of alcohol absorption from AFF diet into the blood via the rumen wall. Yan (1998) found beneficial effects of alcohol-fermented feeds for improving the marbling score of Korean native steers, and Lin (2001) observed that production of total volatile fatty acid and propionate in rumen were affected by supplementation of alcohol-added feed or alcohol-fermented feed, and may improve body weight gain of Korean native steers by decreased protein degradation as well as increased fat synthesis (Lin et al., 2005).
The multiple inputs and outputs of alcohols indicate that alcohol metabolism in ruminants represents a complex system, and the metabolism effects of alcohols by lactating cows is not well understood and has been sparsely described. The present study was undertaken to investigate supplementation of different level of alcohol fermented feed to the TMR on lactating performance, blood metabolites and milk fatty acid profile in Holstein lactating cows.
MATERIALS AND METHODS
Animals and diets
All experimental procedures for this study were approved by the Institutional Animal Care and Use Committee of Kangwon National University, Korea. The study was conducted with forty Holstein lactating cows (106±24 d post-partum; mean±SD) between July 3 and November 11 of 2010. Holstein cows were distributed into groups of four to ensure uniform distribution based on parity, body weight and daily milk yield and assigned to one of four treatments with each containing 10 cows per treatment. Alcohol fermented feed (AFF) was prepared by mixing 45% cracked corn, 45% brewery meal, 5% molasses and 5% live yeast. Then the whole mixture was fermented at 30°C for 48 h. Concentration of alcohol was determined using the alcohol auto-analyzer (Pet-109, Atago, USA). The treatment supplemented with TMR (dry matter basis) as the control (CON), and CON mixed with alcohol-fermented feeds (AFF) at a level of 5%, 10% and 15% of the TMR as T1, T2 and T3, respectively. Lactating Holstein cows were fed 5.0 kg (DM) of TMR per meal twice daily at 08:00 and 18:00 h in equal amounts individually prior to milking. Experimental TMR and AFF feed were supplied by DEAHAN feed Co., LTD in Korea. The feeding level of the TMR was set based on the nutrient requirements for lactating cows (NRC, 2001). The ingredients and chemical composition of the experimental diets are presented in Table 1.
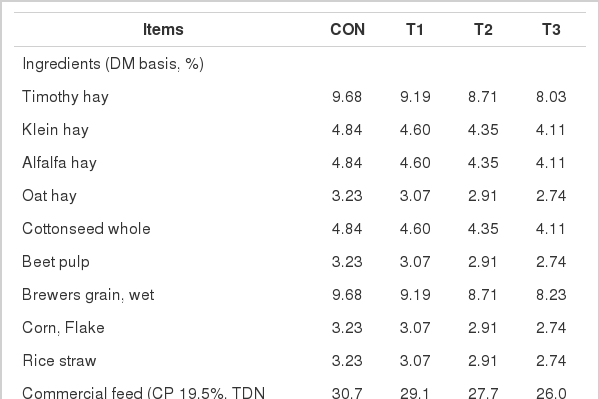
Ingredients and chemical composition of experimental diets
Feed sampling and measurements
Feeds and refusals were collected and amounts were recorded 30 min prior to morning feeding at 08:00 h daily to estimate dry matter intake (DMI). Samples of diets and orts were collected twice per period for chemical analyses. Feed efficiency was calculated by the milk yield per day divided by the feed intake per day of the cow. Feed intake was also calculated by the residual feed deducted from the total supplied feed. AOAC (1995) procedures were used to determine dry matter (DM), crude protein (CP), ether extract (EE), ash, Ca and P. Feed NDF and ADF were analyzed according to the method initially described by Van Soest et al. (1991).
The AFF samples for alcohol determination were obtained after dissolution at 4°C for 30 min followed by 5 X dilutions with distilled water. Then, the samples were centrifuged at 3,000 rpm for 5 min and supernatants were collected. Alcohol concentration in the supernatant was analyzed by glucose analyzer (YSI 2700, USA) with YSI 2,386 alcohol oxidase membrane.
Milk collection and measurements
The Holstein cows were machine-milked (Type MES, No. 843410, Marice Animal Husbandry Machinery Technology Co., Ltd. Wenzhou, China) twice daily at 08:20 and 18:20 h. Milk production was recorded daily, and 100 ml of milk samples were collected once per month throughout the experiment. The a.m. and p.m. milk samples (50 ml/50 ml, v/v) were then combined within cow on every sampling day for analysis of milk composition. Milk total solid, protein, fat, lactose, solid non fat, milk urea nitrogen (MUN) and somatic cell counts (SCC) were analyzed with infrared analysis (Foss Milkoscan FT120; Foss Food Technology Corp., Danmark).
Feed efficiency, 4% FCM and 4% FCM feed efficiency were calculated with the following equations:
Lipids in the milk fat were extracted using Folch’s solution (1957), and methylation of fatty acid (FAME) was prepared according to the method of Lepage and Roy (1986) prior to injection into a gas chromatograph (GC-17A, SHIMADZU, Japan) equipped with a flame ionization detector and a 30 m fused silica capillary column (Supelcowax™ -10, 0.32 mm i.d. Supelco inc., Bellefone, PA, USA). Ultra-high purity He was used as a carrier gas at a flow rate of 1 ml/min. Injector and detector temperatures were 270 and 280°C, respectively. The split ratio of the flame ionization detector was 1:50. The oven temperature was scheduled from 190°C for 5 min and increased to 230°C at a rate of 2°C/min, and was maintained at 230°C for 25 min. A variety of fatty acid standard mixtures (Nu Check Prep; Elysian, MN), were used to identify components and assist in the calculation of response factors. Total cholesterol concentration in milk fat was determined by a gas chromatograph (GC-17A, SHIMADZU, Japan) equipped with a flame ionization detector. The oven temperature was scheduled from 290°C for 5 min and increased to 310°C at a rate of 2°C/min, and was maintained at 310°C for 5 min. The temperatures of the injector and detector were maintained at 270°C and 300°C, respectively. A 30-m fused silica capillary column (HP-1, Crosslinked Methyl Silicone Gum, 0.32 mm i.d. USA) was used. Identification and quantification were carried out by the external standard method following injection of a cholesterol standard (Supelco 47127-U, USA) of known concentration.
Blood collection and analyses
Ten milliliters of blood was taken from the jugular vein into evacuated test tubes (Vacutainer, Becton 93 Dickinson Vacutainer Systems, Rutherford, NJ, USA) containing K3-EDTA 1 h before the morning feeding (08:00 h) once per month throughout the experimental period for analysis of blood metabolites. Blood was centrifuged at 3,000×g for 15 min at 4°C, and serum was stored at −70°C until used for assay. Contents of albumin, blood urea nitrogen (BUN), total protein (TP), creatine, glucose, triglycerides (TG), total cholesterol (TC), HDL-cholesterol (HDL-C), calcium, phosphorus and magnesium in serum were measured using an automated blood serum biochemical analyzer (T Express Plus, USA). The ammonia content of the samples was determined according to the method of Chaney and Marbech (1962) whereas lactic acid content was analyzed by UV spectrophotometer (Hitachi, Japan). Serum gamma gluctamyl transpeptidas (GGT), glutamate oxaloacetate transaminase (GOT) and glutamate pyruvate transaminase (GPT) concentrations of the blood samples were analyzed by Green Cross MS in Korea.
Statistical analysis
Results obtained from the experiment were analyzed by one-way ANOVA for variance using the GLM (General Linear Model) procedure of the SAS (2004) 9.1 Soft Ware Package and authorized synonymy between averages were determined by Duncan’s multiple range test method. Significant differences were accepted if p<0.05.
RESULTS
Dry matter intake, milk yield, milk components and apparent feed efficiency
The DMI and milk yield were not affected by supplementation of AFF (Table 2). Similarly, TMR containing AFF did not alter the contents of total solid, protein, fat, lactose, solid non fat and MUN in milk. An increased 4% FCM in the milk occurred in cows fed T3 diets compared to CON, while T1 and T2 diets decreased 4% FCM and the reduction was in a dose dependent matter. However, apparent feed efficiency of converting feed into energy-corrected milk did not differ among the treatments. TMR containing AFF also decreased milk SCC compared with control treatment.
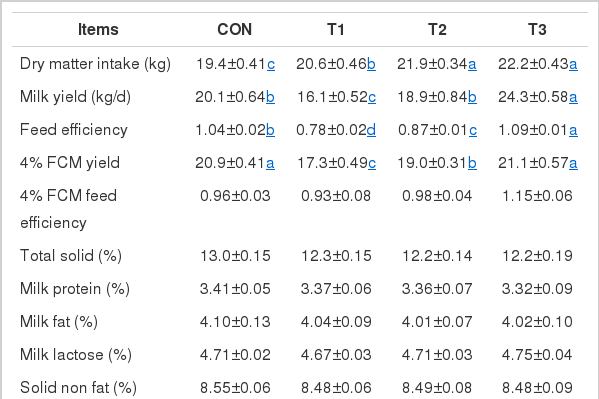
Performance of Holstein lactating cows as influenced by supplementation of alcohol fermented feedstuffs to the TMR
Blood metabolite concentrations
Cows, fed the TMR containing AFF had an increased (p<0.05) concentration of albumin, total protein (TP), ammonia, and high density lipoprotein cholesterol (HDLC) in their serum compared with CON (Table 3), but there was no difference among T1, T2 and T3. In contrast, supplementation with AFF clearly decreased concentration of blood urea nitrogen (BUN) and total cholesterol (TC) in serum compared with CON, and the lowest TC concentration was observed in T3 diets. The concentration of creatinine, total triglyceride (TG), Ca, P and Mg in serum was not influenced by the TMR containing AFF. AFF did not affect the concentration of GGT, GPT and GOT in serum of Holstein cows (Table 3).

Serum metabolites as influenced by supplementation of alcohol fermented feedstuffs to the TMR in Holstein lactating cows
Fatty acid profile and cholesterol concentration in milk fat
The AFF supplementation increased the proportion of C18:1n9 and C18:2n6 compared to CON (Table 4). A decrease (p<0.05) in the content of saturated fatty acid (SFA) for T1, T2 and T3 treatments resulted in an increased unsaturated fatty acid (USFA) to SFA ratio compared to CON. Supplementation with AFF also decreased concentration of cholesterol in milk fat without difference among the T1, T2 and T3 treatments compared to CON (Figure 1).

Fatty acid profile in milk fat (g/100 g FA) of Holstein lactating cows as influenced by supplementation of alcohol fermented feedstuffs to the TMR

Concentration of cholesterol in milk as influenced by supplementation of alcohol fermented feedstuffs to the TMR in Holstein lactating cows. Control: The dairy Holstein cows were fed TMR only, T1: Fed TMR supplemented with alcohol-fermented feed (AFF) at a level of 5% of the TMR, T2: Fed TMR supplemented with AFF at a level of 10% of the TMR, T3: Fed TMR supplemented with AFF at a level of 15% of the TMR. a,b Means within a row with different superscript differ (p<0.05).
DISCUSSION
One of the factors which influence DMI of ruminants is the net energy density of the diet, and alcohol in diets is known to be transformed to VFA by the rumen microorganisms (Andree et al., 1991), and most alcohol is known to be absorbed through the rumen wall (Burning and Yokoyama, 1988). In our current study, an increased DMI and milk yield occurred bin T3, and the increase was dose dependent with an increasing level of AFF. This may due to increased net intake of alcohol, however, milk protein, fat and lactose were not altered whrn the TMR contained AFF. Similar results occurred in lactating cows when fed corn silages with various alcohol concentrations in TMR (Kristensen et al., 2007). However, Randby et al. (1999) found that when cows received ethanol at a daily dose of 600 g in well-fermented grass silage, milk yields decreased slightly, milk fat and protein increased but lactose decreased. Such differences in milk composition secretion could be related to the differences in alcohol feed type and its effects on rumen fermentation metabolism. Furthermore, the highest feed efficiency and 4% FCM in the T3 treatment among the treatments could be due to the large amount of ethanol can be readily utilized by dairy cows (Durix et al., 1991). The lower SCC in milk and an increased milk yield when TMR contained AFF compared with CON treatment in our study indicates that alcohol fermented feed may improve milk quality and milk production.
In ruminants, energy metabolism and blood metabolites can be affected by alcohols, because most alcohols are absorbed through the rumen wall into the blood (Burning and Yokoyama, 1988; Anbarasu et al., 2002; Chen et al., 2002). Change in correlation among the concentrations of albumin, BUN, TP and ammonia after feeding AFF diet implies the possible alteration of protein metabolism in the body. Furthermore, BUN also is a useful indicator of protein metabolism and status in cows (Roseler et al., 1993). In our study, feeding AFF did not affect the concentration of TG in serum. However, an increased TG concentration was observed by Yan et al. (1998) when they examined the effect of alcohol-fermented feeds on blood metabolites in Korea native steers.
In the recent past, dairy products have received adverse publicity in terms of their effect on the blood cholesterol concentration of consumers. They have been perceived to increase blood cholesterol content, which is a risk factor in the aetiology of coronary heart disease. In our study, AFF reduced concentration of cholesterol both in serum and milk. Alteration in correlation between the concentrations of blood cholesterol and milk cholesterol after feeding AFF diet could be due to the possible alteration of lipid metabolism in the body (Pearce, 1996). In contrast, Yan (1998) found higher concentrations of serum cholesterol in Korean native steers when fed alcohol-fermented feeds. Such differences in cholesterol secretion between breeds (i.e. lactating cow and beef steers) could be related to differences in the metabolism of fat (lipoprotein) in the rumen and/or tissues. Because of our limited number of observations, it is not possible to draw definitive conclusions about the level of regulation of cholesterol in milk. The normal concentration of GGT, GPT and GOT enzymes in serum are intracellular and therefore only their higher values have a pathological significance (Simion et al., 2010).
Large amounts of ethanol can be readily utilized by dairy cows (Pradhan and Hemken, 1970). A part of this ethanol is metabolized by rumen bacteria (Moomaw and Hungate, 1963) mainly forming acetate (Czerkawski and Breckenridge, 1972) and, to a lesser extent to the other volatile fatty acids (Pradhan and Hemken, 1970). Supplementation of alcohol increased the proportion of essential fatty acid (C18:2n6, C18:3n9) and unsaturated fatty acid in milk fat. Furthermore, increases in unsaturated fatty acid concentration caused an increase in the ratio of USFA to SFA when the TMR contained AFF. This may suggest that lactating cows fed AFF diet were able to alter the fatty acid profile due to a shift in the rumen fermentation with increase in acetate and mammary de novo fatty acid synthesis.
Feeding a diet supplemented with a moderate level AFF to lactating cows could be a way to alter the feed efficiency and fatty acid profile of milk to increase for the potential human consumer healthy fatty acid without detrimental effects on feed intake and milk production. A substantially decreased cholesterol proportion in milk induced by supplementation with AFF suggests that alcohol fermented feed may improve milk cholesterol level without any negative effects in lactating cows.