Mutations of TYR and MITF Genes are Associated with Plumage Colour Phenotypes in Geese
Article information
Abstract
The polymorphism of microphthalmia-associated transcription factor (MITF) and tyrosinase (TYR) genes have been proposed to play a vital role in coat colour genesis in mammals, but their role remains ambiguous in geese at best. Here, we cloned and sequenced 1,397 bp coding region of MITF gene and a 588 bp fragment of TYR exon 1 for polymorphism analysis among 157 domestic geese showing three types of plumage colour. We detected a total of three SNPs (c.280T>C, c.345G>A, and c.369G>A) in TYR and six haplotypes (H1–H6). Among them, haplotypes H1, H2, H3, and H5 were significantly associated with white plumage trait of Zhedong White Geese. However, only diplotype H1H1 and H3H5 were significantly associated with white plumage trait of Zhedong White Geese (p<0.01). We only detected one SNP (c.1109C>T) for MITF gene and found that genotype CT and TT were significantly associated with white plumage trait of Zhedong White Geese. Briefly, our study suggested an association between polymorphisms of TYR and MITF genes and the plumage colour trait in domestic geese.
INTRODUCTION
The plumage, skin and eye colour of birds are mainly determined by the relative content and distribution of eumelanin and phaeomelanin, which is produced by melanocytes present in the epithelium, iris, and feather follicles of birds. The eumelanin is brownish-black and the phaeomelanin range of colour is from yellow to reddish-brown (Prota, 1980). The process of melanin biosynthesis is triggered by the binding of α-melanocyte stimulating hormone (α-MSH) to its receptor, melanocortin-receptor 1 (MC1R). The subsequent increasing level of cyclic AMP (cAMP) up-regulates the transcription of microphthalmia-associated transcription factor (MITF). Afterwards, MITF activates the transcription of tyrosinase, which catalyzes the first step of melanogenesis (Smyth, 1990; Scherer and Kumar, 2010). The tyrosinase, encoded by tyrosinase (TYR) gene, is known to be the rate-limiting enzyme affecting the production of melanin pigment (Sánchez-Ferrer et al., 1995), which oxidates tyrosine to dihydroxyphenylalanine (DOPA) (Lerner and Fitzpatrick, 1950) and determines which type of melanin (eumelanin or phaeomelanin) could be synthesized (Ito et al., 2000). After the report of (Oetting et al., 1985) that the C locus is the structural locus for tyrosinase in the fowl, the TYR gene was first cloned from chicken (Mochii et al., 1992). Previous studies have shown that mutations of the TYR gene are associated with albinism phenotype in many vertebrates, such as human (Fukai et al., 1995; Oetting, 2000), mouse (Yokoyama et al., 1990), rat (Blaszczyk et al., 2005), cat (Imes et al., 2006), rabbits (Aigner et al., 2000), cattle (Schmutz et al., 2004), chicken (Tobita-Teramoto et al., 2000; Sato et al., 2007; Liu et al., 2010).
MITF is a transcription factor and belongs to the basic-helix-loop-helix-leucine zipper (bHLHZip) protein family. Studies suggested that it regulates the melanogenesis through binding to a highly conserved M-box (GTCATGTGCT) and E-box (TCATGTG) motif upstream of the tyrosinase promoter, which strongly stimulates the tyrosinase promoter activities and promotes tyrosinase expression (Yasumoto et al., 1994; Bertolotto et al., 1996, 1998). Besides, it is also involved in the development of melanocytes, retinal pigment epithelium, mast cell and osteoclasts (Hodgkinson et al., 1993; Steingrímsson et al., 1994; Moore, 1995). Mutations of the MITF gene that have been described in several vertebrate species lead to coat colour dilutions, white spotting, or complete loss of pigmentation in mouse (Hodgkinson et al., 1993; Hughes et al., 1993; Yasumoto et al., 1998), cause Waardenburg syndrome (Variant, 1994) and Tietz syndrome (Amiel et al., 1998; Smith et al., 2000) in human with the characteristics of hearing loss and depigmentation of the hair and skin. Subsequent studies also reported that mutations in MITF are associated with white spotting coat colour in dogs (Andersson, 2009; Schmutz et al., 2009) and white plumage in Japanese quail (Minvielle et al., 2010).
Nowadays, coat color is considered an important economic trait that caters to consumer preference. Geese are valued as much for their feathers when processed into clothing for warmth and fashion shows. However, little is known about the genetic basis of feather color. As a qualitative trait, the plumage colour of poultry is determined by the relative content and distribution of eumelanin and phaeomelanin produced in melanocytes. The melanin synthesis is controlled by the sequential expression and interactions of genes (Scherer and Kumar, 2010). Taken together, the aforementioned reports have shown that the polymorphisms of TYR and MITF genes are the key factors for melanin synthesis and albino phenotype expression in mammals. However, the potential association between MITF and TYR genes and plumage colour phenotypes in geese has not been validated. The aim of the current study is to detect SNPs in MITF and TYR genes and explore their possible association with plumage colour phenotypes in goose.
MATERIALS AND METHODS
Animals
Blood samples from 157 geese with three plumage colour phenotypes were collected within two breeds, including 41 gray plumage Landes (completely gray feathers at dorsum and breast), 42 spotted feather Landes (near black feathers at dorsum and breast), 74 Zhedong White Geese (pure white plumage). All samples were gathered from the Research Poultry Farm Veterinary Research Institute of Shanghai Academy of Agricultural Sciences where special staff raised the birds at the same level of nutrition and management in a poultry house. Meanwhile, skin hair-follicle tissues were collected from Sichuan white geese for RNA extraction and cDNA synthesis to clone the CDS (coding sequence) of MITF gene.
DNA extraction, sequencing and genotyping of MITF and TYR gene
The genomic DNA was prepared by a standard phenol/chloroform method to proceed the PCR amplification and sequencing of TYR and MITF genes to detect and analyze the sequence mutations. One pair of primers were designed by Oligo 6.0 and Primer 5.0 software based on the conserved sequence of exon 1 of TYR gene in Gallus gallus (NM_2046160, L46805), Meleagris gallopavo (XM_003203534) and Cotunix cotunix (AB024279). Another pair of primers were designed for sequencing of a partial coding sequence of MITF (Minvielle et al., 2010) (Table 1). A total volume of 50 μL PCR reaction mixture was used containing 25 μL 2×Taq Master Mix (TaKaRa Biotech Co. Ltd., Dalian, China), 100 ng genomic DNA, 2.5 μL of each primers and 17 μL ddH2O. The 35 PCR cycles were carried out as follows: 94°C for 30 s, the respective annealing temperature of the primer pair for 45 s, 72°C for 1 min, preceded by a denaturizing step of 5 min at 94°C and followed by an extension step of 10 min at 72°C. Subsequently, the PCR products of all 157 samples of TYR and MITF genes were purified with centrifugal columnar agarose gel DNA extraction kit (TianGen Biotech Co. Ltd., Beijing, China) and sequenced directly on ABI Prism 3700 DNA sequencer (Shanghai Biotech Co. Ltd., Shanghai, China).

Primers for MITF and TYR genotyping and cloning of MITF
RNA isolation, cDNA synthesis and CDS clone of MITF
Total RNA was isolated from skin hair-follicle tissues of Sichuan white goose using Trizol reagent (TakaRa Biotech Co. Ltd., Dalian, China). The quality of RNA was determined by the A260/280 absorbance ratio (1.6–1.8) and the integrity of the 18S and 28S rRNA bands was checked on 1% formaldehyde agarose gel. Isolated RNAs were treated with 8 μL DNase (Fermentas) at 37°C for 20 min and stored at −80°C. The cDNAs were synthesized using the ImProm-II Reverse Transcription System (TakaRa Biotech Co. Ltd., Dalian, China) according to the manufacturer’s recommendation. The reaction was performed in a volume of 10 μL, containing 5×PrimerScript Buffer, 10 mM of each dNTPs, 40 U/μL RNase Inhibitor, 2.5 μM oligo-dT Primer. The reverse transcription was maintained at 37°C for 15 min, and 85°C for 5 s. The cDNA samples were stored at −80°C.
The primers for cloning MITF gene were designed by Oligo 6.0 and Primer 5.0 software according to the conserved mRNA sequence of MITF in Gallus gallus (NM_205029), Meleagris gallopavo (XM_003210180), Cotunix cotunix (AB005229) and Taeniopygia guttata (XM_002193131) (Table 1). PCR amplification was performed in a final volume of 50 μL containing 25 μL 2× Taq Master Mix (TaKaRa Biotech Co. Ltd., Dalian, China), 4 μL cDNA template, 1.5 μL of each primers and 19.5 μL ddH2O with the following cycling parameters: one cycle of 5 min at 94°C, 35 cycles of 30 s at 94°C, 45 s, 45 s at the respective annealing temperature of each primer pair, 1 min at 72°C, and a final extension for 10 min at 72°C. Purified PCR products were cloned into pMD 18-T simple vector (TakaRa Biotechnology Co. Ltd., Dalian, China). The authenticity of the amplified fragment was verified by sequencing.
Data analysis
All sequences were spliced and aligned by DNAstar package (DNASTAR Inc., Madison, WI, USA). We used MEGA 4.0 to export sequence variations. Haplotypes of the TYR gene were deduced by using the PHASE 2.0 program (Stephens et al., 2001). The potential association between the TYR and MITF alleles and plumage colours were evaluated by chi-square test for independence and which was performed on SAS V8.1 (SAS Institute Inc. Cary, NC, USA).
RESULTS AND DISCUSSION
We obtained 588 bp fragments of exon 1 of TYR gene and 1,407 bp coding fragments of MITF gene in this study and the sequence data was deposited in GenBank under accession numbers of KF020633 and KF020632, respectively. Three SNPs (c.280T>C, c.345G>A and c.369G>A) were detected from TYR gene, among which c.345G>A and c.369G>A were only found in Zhedong White Geese and the c.280T>C was found in Zhedong White Geese and spotted plumage colour Landes, no polymorphism was detected in grey plumage colour Landes (Table 2). However, only one SNP (c.1109C>T) was detected in the MITF gene exclusively in Zhedong White Geese containing two alleles and deducing three genotypes (CC, CT, TT). All of these SNPs were synonymous and newly reported. As is known, the preference of tRNA binding degenerate codons generated by synonymous mutations is not random in the process of gene translation and the synonymous mutation would affect and reduce the translation efficiency of gene, and subsequently affect the protein folding and function (Kimchi-Sarfaty et al., 2007; Komar, 2007). However, the genotypic milieu was not the reason for the occurrence of those SNPs, and the hypothesis of tRNA binding preference might explain why those synonymous mutations were only distributed in specific groups.

The genetic polymorphisms in TYR gene and MITF gene
The MITF genotypes CT and TT were significantly associated with the white plumage trait of Zhedong White Geese (p<0.05) which further verified that the mutation c.1109C>T affected the plumage color (Tables 2 and 3). While in most dogs an interspersed nucleotide element (SINE) insertion 3167 bp before the start codon of exon 1M in MITF plays an important role in coat color determination. Dogs homozygous for SINE had white markings that covered most of the body, while the heterozygous ones were solid colored or had minor white (Schmutz et al., 2009). The distinct patterns in geese and dogs provides a glimpse of the MITF association with coat color.

The genotype distribution of TYR gene and the MITF gene
In addition, six haplotypes (H1 to H6) were obtained based on the three SNPs (c.280T>C, c.345G>A, and c.369G>A) of the TYR gene (Table 4), and the corresponding diplotypes are displayed in Table 5. The results showed that the haplotypes H1, H2, H4, and H5 were only found in Zhedong White Geese. Particularly, haplotype H6 occurred in Zhedong White Geese and spotted Landes with the same frequency. Association analysis demonstrated that haplotypes H1, H2, H3, and H5 were significantly associated with the white plumage trait of Zhedong White Geese, whereas the H2 and H3 were significantly associated with gray and spotted plumage trait of Landes (p<0.01). Furthermore, the diplotypes were only distributed in Zhedong White Geese, besides diplotype H3H3. Association analysis showed that diplotypes H1H1 and H3H5 were significantly associated with white plumage trait of Zhedong White Geese (p<0.01). We can infer that the difference diplotypes of TYR exon 1 effect the plumage color determination. While in cats, three mutations (c.216G>T, c.450C>T, c.679G>T) were identified in exon 1 of TYR with one nonsynonymous and two synonymous mutations which showed strong association with albino locus (Schmidt-Küntzel et al., 2005). In Himalayan mice, strong association also been proved between a nonsynonymous substitutions in exon 4 of TYR and the albino locus (Kwon et al., 1989). Through different species, we can speculate that nucleotide mutation plays a crucial role in TYR on coat color determination.

Distribution of the TYR gene haplotypes in three goose populations
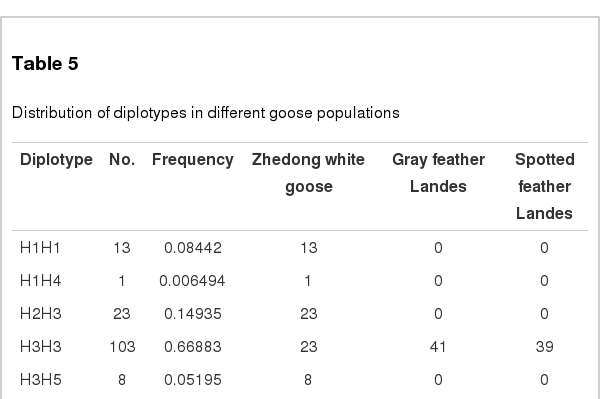
Distribution of diplotypes in different goose populations
As a complex trait, coat color is determined by a complex pathway system and multiple interactively patterns, further investigation would be helpful to further elucidate the molecular mechanism of plumage color formation. On the basis of our result in this study, we can speculated that, in geese, TYR and MITF genes play a vitally important role in plumage color determination.
ACKNOWLEDGEMENTS
This work was funded by the National Natural Science Foundation of China (31172181) and by Sichuan Province (11TD007; 2011JTD0032).