Effects of Lactation Stage and Individual Performance on Milk cis-9, trans-11 Conjugated Linoleic Acids Content in Dairy Cows
Article information
Abstract
The goal of this study was to evaluate the effects of lactation stage and individual performance on milk cis-9, trans-11 conjugated linoleic acid (CLA) content in dairy cows. In experiment 1, the milk cis-9, trans-11 CLA content from dairy cows in early (0.33±0.014%), middle (0.37±0.010%), and late stages (0.44±0.020%) showed significant differences (p<0.05); and the individual contents of the major fatty acids, especially cis-9, trans-11 CLA in cows of the same lactation were also variable. In the second experiment design as a validation test, our results once again showed that the individual contents of cis-9, trans-11 CLA were various, and a difference of about 2-fold (0.55% vs 0.95%) was observed, although the animals were offered same diet. These data demonstrated that lactation stage and individual performance have considerable effects on milk cis-9, trans-11 CLA contents.
INTRODUCTION
Conjugated linoleic acid (CLA) is a group of isomers that has beneficial health effects in humans and animals (Bauman et al., 2000). Of the various potential isomers, cis-9, trans-11 CLA accounts for 82 to 97% and 57 to 85% of the total CLA content in milk and beef, respectively (Dhiman et al., 2005). This specific isomer exhibits several health benefits that are not shown by other CLA isomers (Wang and Lee, 2012) thereby leading to heightened interest in the manufacture of cis-9, trans-11 CLA-fortified products (Dhiman et al., 2005; Rodríguez-Alcalá and Fontecha, 2007). The majority of the cis-9, trans-11 CLA found in milk is synthesized in mammary glands, in a process catalysed by stearoyl-CoA desaturase 1 (SCD1), while the remaining part is derived from the rumen (Bauman et al., 2000; Griinari et al., 2000; Wang and Lee, 2012). SCD1 is one of the two identified SCD isoforms in the bovine (Paton and Ntambi, 2009). It is an enzyme that predominantly exists in the endoplasmic reticulum (ER) membrane of liver, mammary gland, adipose tissue, etc. (Wang and Lee, 2012). SCD5 is another SCD isoform which is highly expressed in the brain (Lengi and Corl, 2007) and it is also expressed in the bovine mammary tissue (Jacobs, 2011).
Factors that regulate the SCD1 activity or rumen fermentation conditions can alter the milk content of cis-9, trans-11 CLA. The effects of breed, diet, feeding strategies and seasonal variations have been reported (Dhiman et al., 2005; Slots et al., 2008). However, the effects of lactation stage and individual variation among dairy cows on the cis-9, trans-11 CLA content are still not well understood. Therefore, in this study, two experiments were conducted in order to evaluate the effects of lactation stage and individual performance on milk cis-9, trans-11 CLA content in dairy cows.
MATERIALS AND METHODS
Cows and milk samples
Two experiments were performed in this study. All experimental procedures were in accordance with the “Guidelines for the Care and Use of Experimental Animals of Pusan National University”. Experiment 1 employed thirty-one multiparous Holstein cows fed the same total mixed ration (TMR) (Tables 1 and 2). Experiment 2 used seventeen multiparous Holstein cows (589±29.6 kg of body weight) in the middle lactation stage (142.5±29.65 d) and fed the same diet. The ratio of roughage to concentrates was about 0.77:1 (dry matter basis); 10 kg of oat hay (dry matter 91.5%, crude protein 1.2%, ether extract 2.3%, neutral detergent fiber 58.3%, acid detergent fiber 36.7%, and ash 8.3%) and 13 kg of concentrate were fed daily. The ingredient (% DM), proximate (% DM), and fatty acid (% total fatty acids) compositions of the concentrate fed to the dairy cows in experiment 2 are shown in Table 3. Water was available at all time for all the animals. Cows were milked twice daily (12-h milking interval). The morning milk was collected from animals individually after machine milking from the four quarters. The bulk tank somatic cell counts (SCC) were monitored and the values were less than 100,000 cells/ml. The milk samples were stored at −20°C till milk fat analysis.

Design of experiment 1

Ingredients (kg, wet matter) and proximate (% dry matter) composition of the TMR fed to dairy cows (Experiment 1)

Lipid analysis
Briefly, total milk lipids were extracted with 20 ml chloroform/methanol (2:1, v/v) from 6 ml of milk. The extracted lipids were then converted into fatty acid methyl esters with 14% (w/v) boron trifluoride-methanol (B1252; Sigma-Aldrich Corp., St. Louis, MO, USA) according to a previously published method (Folch et al., 1957). The fatty acid methyl esters were then injected into a gas chromatograph (GC) (Agilent 7890A GC system, Agilent Technologies, USA) using a system equipped with a 7863 series auto-sampler, 7683B series injector, and flame ionization detector (FID). A SP™-2560 fused silica capillary column (100 m×0.25 mm, inner diameter with 0.2-μm film thickness; Supelco Inc., Bellefonte, PA, USA) was used and the oven temperature was programmed to increase from 70 to 225°C at a rate of 5°C per min to 100°C, held for 2 min, increased 10°C per min to 175°C, held for 40 min, increased 5°C per min to 225°C, and held for 40 min. Front inlet and FID temperatures were maintained at 255°C and 260°C, respectively. The hydrogen flow rate to the detector was 55 ml/min, the airflow rate was 400 ml/min, and the make-up gas flow rate was 20 ml/min. The split ratio was 30:1. Peaks were routinely identified by comparing the retention time with those of authentic standards including Supelco® 37 Component Fatty acid methyl esters (FAME) Mix (47885-U), trans-11-octadecenoic methyl ester (46905-U; Sigma-Aldrich Corp.) and cis-9, trans-11 CLA (1255; Matreya LLC, Pleasant Gap, PA, USA). The percentage of individual fatty acid was calculated as the ratio of individual area to that of total identified fatty acids.
Statistical analysis
Data were presented as mean±SE and analyzed using a one-way analysis of variance (one-way ANOVA) (SPSS Inc., Chicago, USA). In all cases, significant differences were accepted if p<0.05. The relationship between cis-9, trans-11 CLA content and desaturation indexes were analyzed using the correlation procedures (PROC CORR) of SAS (SAS, 2000).
RESULTS AND DISCUSSION
Effects of lactation stage on milk cis-9, trans-11 CLA content
Previous studies have shown that lactation stage significantly contributes to variation in the milk fat profile (Kay et al., 2005; Stoop et al., 2009). Normally, linoleic acid can be isomerized into cis-9, trans-11 CLA in the rumen, with the majority being further hydrogenated into trans-11 18:1, and then potentially saturated into stearic acid (Wang and Lee, 2012). In experiment 1, both the linoleic acid and stearic acid levels significantly decreased from the early stage of lactation (before 100 d) to the late stage (after 200 d) (p = 0.032 and p = 0.022, respectively; Table 4). Because the animals were offered the same diet, the decreased linoleic acid and stearic acid levels in milk may suggest increased activity of rumen microorganisms and more rumen output of trans-11 18:1 by dairy cows in the late stage of lactation.

Differences in major milk fatty acids (%) from dairy cows in Experiment 1
The milk trans-C18:1 content of dairy cows in middle stage of lactation was significantly lower than that in early stage of lactation (p = 0.003) whereas no significant difference was found between the late stage and early stage (p = 0.087) (Table 4). Three desaturation indexes were calculated representing the ratio of the Δ9-desaturase product to the sum of the Δ9-desaturase product and substrate (Kelsey et al., 2003) (Table 4). Although not statistically significant, the desaturation indexes for cis-9 C14:1 and cis-9 C18:1 in the late stage of lactation were all a little higher than that in the early stage of lactation (p = 0.116 and p = 0.609, respectively; Table 4) and these results may indirectly imply a higher activity of Δ9-desaturase of the mammary glands of animals in the late stage of lactation. However, no difference was found for the desaturation index for cis-9 C16:1. As we have speculated, the cis-9, trans-11 CLA continually increased from the early stage of lactation to the late stage of lactation (0.33% vs 0.44%; Figure 1). A similar result was also found in the milk of Canadian Jersey cows, with the cis-9, trans-11 CLA content in the late stage (after 200 d) being significantly higher than during early lactation (before 100 d) (Kgwatalala et al., 2009). In a dairy ewe research, greatly increased cis-9, trans-11 CLA content was observed in Transylvania Merino sheep (2.03 g vs 2.70 g/100 g FAME), Tsigay sheep (2.05 g vs 2.60 g/100 g FAME) and Turcana sheep (2.36 g vs 3.64 g/100 g FAME) from 2 wks to 14 wks of lactation (Mierlita et al., 2011). However, in another report it was said that the cis-9, trans-11 CLA content of bovine milk and Δ9-desaturase activity in the mammary gland are independent of stage of lactation under normal conditions (Lock et al., 2005).
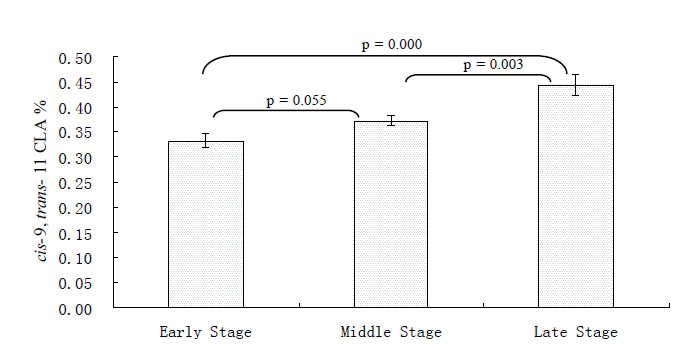
cis-9, trans-11 CLA content in milk from dairy cows at different lactation stages (Experiment 1).
Effects of individual performance on milk cis-9, trans-11 CLA content
The individual contents of the major fatty acids, especially cis-9, trans-11 CLA in cows of the same lactation were found variable in experiment 1. In milk, cis-9, trans-11 CLA contents were 0.27 to 0.38%, 0.30 to 0.42%, and 0.36 to 0.57% in the early stage, middle stage and later stage, respectively (Table 4). These data suggested large individual differences in the cis-9, trans-11 CLA synthesis ability of cows offered the same diet and this may be related to both rumen biohydrogenation and Δ9-desaturase activity in the mammary gland (Lock and Garnsworthy, 2002; Peterson et al., 2002).
In order to confirm if individual performance really had effect on milk cis-9, trans-11 CLA content, experiment 2 was designed as a validation test. More animals in middle lactation were used in this experiment (17 multiparous Holstein cows) compared with that in experiment 1 (11 multiparous Holstein cows). Our results once again showed that the individual contents of cis-9, trans-11 CLA were various, and a difference of about 2-fold (0.55% vs 0.95%) was observed although the animals were offered same diet (Figure 2). Corresponding differences were also evident in the desaturation indexes for cis-9 C14:1 (0.069 vs 0.137) and cis-9 C16:1 (0.036 vs 0.064) (Table 5). These substantial variations in the desaturation indexes may demonstrate a huge diversity of the Δ9-desaturase activity in mammary glands of dairy cows. A positive correlation (R2 = 0.558) existed between cis-9, trans-11 CLA content and the desaturation index for cis-9 C14:1 (Figure 3). However, no obvious correlation was found between cis-9, trans-11 CLA content and the desaturation index for cis-9 C16:1 (R2 = 0.087) or cis-9 C18:1 (R2 = 0.218) (Figure 3). Moreover, large variations were found in the milk content of linoleic acid (2.47% to 5.32%) and linolenic acid (0.34% to 0.68%) further suggesting considerable differences in the rumen environments of these animals. In addition, various individual cis-9, trans-11 CLA contents were also found in two crossbreed dairy ewes including Karakachan× Blackhead Pleven (13.5 to 28.8 mg/g fat) and Tsigay× Blackhead Pleven (13.7 to 43.6 mg/g fat) (Mihailova and Odjakova, 2011). Lastly, two additional aspects were again confirmed from the results of these 2 experiments (Table 4; Figure 3). First, higher desaturation indexes correspond to higher cis-9, trans-11 CLA content and second, the desaturation index of cis-9 C14:1 appears to be a much more reliable indicator than that of cis-9 C16:1 or cis-9 C18:1.

cis-9, trans-11 CLA content of milk from individual multiparous dairy cows in the middle lactation stage (Experiment 2).

Important parameters and major milk fatty acids composition (%) from dairy cows in Experiment 2
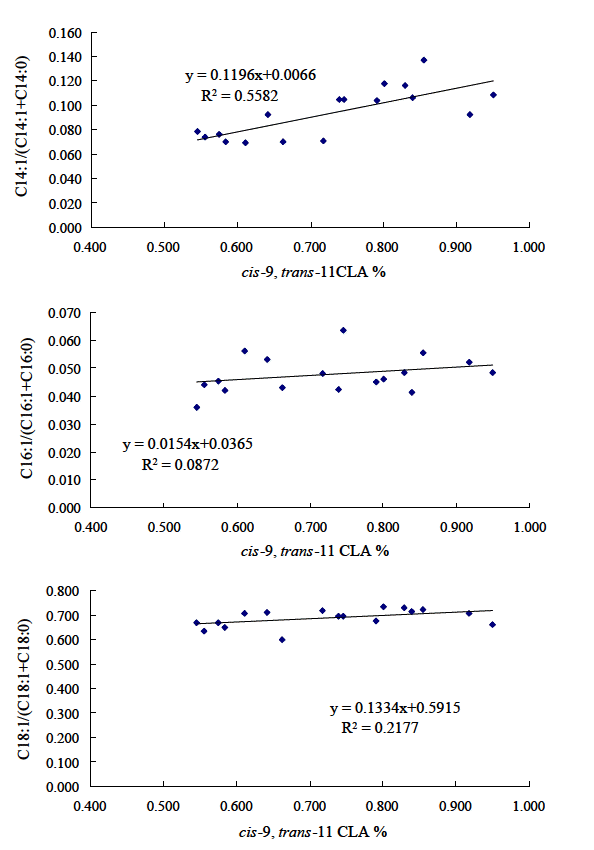
Correlation between cis-9, trans-11 CLA content and desaturation indexes (Experiment 2).
In conclusion, our results demonstrated that lactation stage and individual performance have considerable effects on the milk fat profile in dairy cows, especially in the content of cis-9, trans-11 CLA. Furthermore, the basis for the variation of cis-9, trans-11 CLA content at the individual level and during lactation may be related to the rumen output of trans-11 18:1 and to the activity of Δ9-desaturase in the mammary glands.
ACKNOWLEDGEMENTS
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (Project No. 010-0006707).