Influence of Protein and Energy Level in Finishing Diets for Feedlot Hair Lambs: Growth Performance, Dietary Energetics and Carcass Characteristics
Article information
Abstract
Forty-eight Pelibuey×Katahdin male intact lambs (23.87±2.84 kg) were used in an 84-d feeding trial, with six pens per treatment in a 2×2 factorial design arrangement. The aim of the study was to evaluate the interaction of two dietary energy levels (3.05 and 2.83 Mcal/kg ME) and two dietary protein levels (17.5% and 14.5%) on growth performance, dietary energetics and carcass traits. The dietary treatments used were: i) High protein-high energy (HP-HE); ii) High protein-low energy (HP-LE); iii) Low protein-high energy (LP-HE), and iv) Low protein-low energy (LP-LE). With a high-energy level, dry matter intake (DMI) values were 6.1% lower in the low-protein diets, while with low-energy, the DMI values did not differ between the dietary protein levels. Energy levels did not influence the final weight and average daily gain (ADG), but resulted in lower DMI values and higher gain efficiencies. No effects of protein level were detected on growth performance. The observed dietary net energy (NE) ratio and observed DMI were closer than expected in all treatments and were not affected by the different treatments. There was an interaction (p<0.03) between energy and protein level for kidney-pelvic and heart fat (KPH), KPH was higher in lambs fed high energy and high protein diet but not in high energy and low protein diet. The KPH was increased (20.2%, p = 0.01) in high-energy diets, while fat thickness was increased (21.7%, p = 0.02) in high-protein diets. Therefore, it is concluded that dietary energy levels play a more important role in feed efficiency than protein levels in finishing lambs with a high-energy diet (>2.80 Mcal/kg ME). Providing a level of protein above 14.5% does not improves growth-performance, dietary energetics or carcass dressing percentage.
INTRODUCTION
The amount of tissues deposited as carcass components is largely determined by the level of protein intake and the energy available for retention in muscle (Baracos, 2005). However, the relationship between energy and protein concentration within the diet is not completely elucidated in lambs. It is thought that, in low-to-moderate energy diets (<2.50 Mcal/kg ME), the extra-caloric effect of the increased metabolizable protein intake due to increased protein levels in the diet is likely to be shown by a greater dry matter intake (DMI), a higher average daily gain (ADG) and an increase in the performance of lambs (Muwalla et al., 1998). Moreover, some reports indicate that lambs fed high-energy diets (>2.80 Mcal/kg ME) with an increased protein level above 14% have not shown an advantage in growth performance (Beauchemin et al., 1995), but the body composition of lambs can be modified (NRC, 2007). However, the latter can be affected by the potential for growth (breed), the initial weight at fattening, the number of days on feed and/or the weight at harvest (Cannas et al., 2004). Although breeders have introduced Dorper, Katahdin and Saint Croix sheep breeds to the country of México in recent years, Pelibuey lamb breeds and their crosses continue to be the most representative genotype, and have made an important contribution to the persistence of Mexican lambs (Partida and Martínez, 2010). Pelibuey lambs and their crosses are usually offered growing-finished diets that contain between 2.8 and 3.0 Mcal/kg of ME and 17% to 18% of crude protein. At fattening, hairy lambs and their crosses have shown some differences in growth rate and carcass composition in respect to other breeds (Partida and Martínez, 2009; Ruiz-Nuño et al., 2009). However, very little information is presently available regarding the relationship between protein levels in high energy diets on growth performance and carcass characteristics in finishing hairy lambs.
The objective of this experiment was to evaluate the growth performance, dietary energetics, and carcass characteristics in lambs fed two energy concentrations in a finishing diet (3.05 and 2.83 Mcal/kg ME) at two protein levels (17.5% and 14.5% CP).
MATERIALS AND METHODS
This trial was conducted at Universidad Autónoma de Sinaloa Feedlot Lamb Research Unit, located in Culiacán, México (24° 46′ 13″ N and 107° 21′ 14″W). Culiacán is about 55 m above sea level, and has a tropical climate. All procedures involving animal care and management were in accordance with and approved by the Universidad Autónoma de Sinaloa Animal Use and Care Committee.
Sixty Pelibuey×Katahdin lambs were received at the research facility 7 weeks before the start of the trial. Upon arrival, lambs were weighed, treated for endoparasites (Tasasel 5%, Fort Dodge, Animal Health, México), and injected with 1×106 IU of vitamin A (Synt-ADE, FortDodge, Animal Health, México). During a 35-d adaptation period all lambs received a diet containing (DM basis, g/kg): alfalfa hay, 150; ground grain corn, 555; soybean meal, 200; cane molasses, 70; mineral supplement, 25. Following a 7 wk receiving-evaluation period, 48 lambs, initial weight (IW) (23.8±2.84 kg), were selected from the original group of 60 lambs for use in the study, based on uniformity of weight and general condition. Upon initiation of the trial, lambs were weighed before their morning meal using an electronic scale (TORREY TIL/S: 107-2691, TOR REY electronics Inc, Houston TX, USA), and were assigned within 6 weight groupings to 24 pens (4 pens/treatment), with 2 lambs/pen. Pens were 6 m2 with overhead shade, automatic waterers and 1 m fence-line feed bunks. To evaluate the interaction of two dietary energy levels (3.05 and 2.83 Mcal/kg ME) and two dietary protein levels (17.5% and 14.5%), lamb groups were fed one of four of the next experimental diets over 84 days (Table 1) in a 2×2 factorial design arrangement: i) High protein-high energy diet (HP-HE); ii) High protein-low energy diet (HP-LE); iii) Low protein-high energy diets (LP-HE), and iv) Low protein-low energy diets (LP-LE). Lambs were weighed before the morning meal on d 1 and 84 (harvest). Lambs were fed twice daily at 09:00 and 15:00 h in a 30:70 proportion (as a feed basis). Feed bunks were evaluated visually before feeding between 08:40 and 08:50 h each morning to determine the quantity of feed remaining from the previous day. Daily feed allotments to each pen was adjusted to allow for minimal (<5%) feed accumulation. Adjustments (increase or decrease) in daily feed delivery were allotted to the afternoon feeding. Feed samples were collected on a daily basis for DM analysis (forced-air oven; AOAC, 2000). Assuming that the DMI intake is related to energy requirements and dietary NEm, it is expected that the DMI can be estimated from average ADG and LW values according to the following equation:
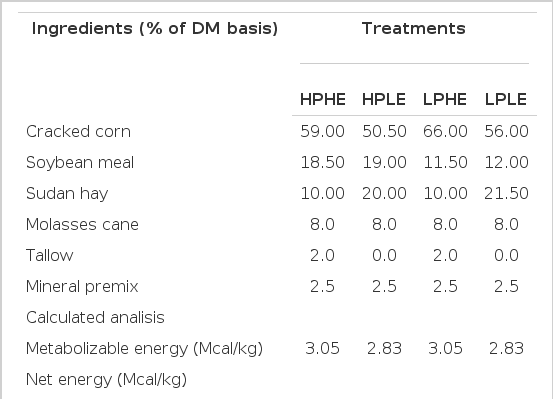
Ingredients and composition of the experimental diets (DM)
Hot carcass weights (HCW) were obtained for all lambs at the time of slaughter. Because feed and water were not withdrawn for 12 h before weighing and slaughtering, the initial and final weights were reduced (pencil shrink) by 4% to account for digestive tract fill (Cannas et al., 2004). After carcasses were chilled for 24 h at 4°C the carcass cold weight (CCW) was recorded and the following measurements were obtained: i) Fat thickness over the 12th rib, perpendicular to the Longissimus dorsi (LM) with a Vernier Caliper (Digital King, foot of 6″, Mexico City), ii) LM area, between 12 to 13th rib was measured using a plastic grid and iii) kidney, pelvic and heart fat (KPH), which was removed from the hind saddle and weighed and reported as a percentage of carcass weight (USDA, 1982). Each carcass was split in half. The right side was merchandised, while the left side was fabricated into wholesale cuts, without trimming, according to the North American Meat Processors Association guidelines (NAMP, 1997), with each cut dissected into the lean components, bone and fat. Components were weighed and the total weights of muscle, fat and bone were expressed in percentages in relation to the cold carcass weight.
Performance (gain, feed efficiency, dietary energetics) and carcass data were analyzed using a randomized complete block design using the MIXED procedure of the SAS Institute (2004) for a randomized complete block design. This design was arranged as a 2×2 factorial experiment, with two levels of energy (3.05 and 2.83 Mcal/kg ME) and two levels of protein (17.5% and 14.5%), using pens as an experimental unit. The fixed effect consisted of treatment, and the random effect was the pen. Carcass composition data were analyzed using the MIXED model procedures (SAS Institute, 2004), including the fixed main effect of treatment and the random effect of individual carcass. Treatment effects were tested as follows: i) Energy level, ii) protein level, and iii) energy×protein interaction. Contrasts were considered significant when the p-value was ≤0.05, and tendencies were identified when the p-value was between 0.05 and 0.10.
RESULTS AND DISCUSSION
The treatments effects on the growth performance of finishing lambs are shown in Table 2. There was an nteraction (p = 0.03) between energy levels and protein levels on the DMI (as percentage of BW). With the high-energy diet, the DMI values were 6.1% lower than with low-protein diets, while with the low-energy diets, the DMI did not differ between proteins levels. In the same manner, Fluharty and McClure (1997) observed lower (12.4%) intakes in lambs when protein levels of 14.3% and 18.9% were compared in a similar high-energy diet (2.80 Mcal NEm/kg) as used in the present study.

Influence of protein and energy level on growth performance of feedlot hair lambs
Lambs fed a high-energy diet had a lower DM intake (13.4%) than the low-energy diet group. This finding is in agreement with the principle that feed intake is regulated by dietary energy density in ruminants. ME intake, rather than physical fill, appeared to be the dominant factor influencing the DMI for the diets investigated. Lu and Potchoiba (1990), observed a curvilinear response when comparing 3 levels of energy (2.46, 2.77, and 3.05 Mcal/kg ME DM) in diets. In a study by Lu and Potchoiba (1990), the maximal DMI was observed in the medium-energy diet (2.77 Mcal/kg ME), which was similar to the energy concentration of 2.83 Mcal/kg ME tested in the present study. The energy level did not influence ADG (p≥0.23), however, the lambs fed high-energy diets showed an increase in gain to feed (8.1%) as a consequence of differences in DM intake. Increases in feed efficiency have been a common response when comparing high-energy and low-energy diets (NRC, 2007; Kioumarzi et al., 2008; Adbel-Basset, 2009). However, the effect of increased dietary energy levels on ADG has been less consistent. In some instances (Craddock et al., 1974; Lu and Potchoiba, 1990; García et al., 2003), increasing the energy level had no effect on the ADG, whereas in others (Kioumarzi et al., 2008; Adbel-Basset, 2009), an increase in energy level markedly increased the ADG. In most cases, when growth performance was compared among dietary energy concentrations below 2.6 Mcal/kg ME, the ADG and feed efficiency increased as energy increased. However when the energy concentration is above of 2.8 Mcal/kg ME, feed efficiency increases with increased energy level without any differences in the ADG (Lu and Potchoiba, 1990; García et al., 2003). The latter could be partially explained by the inverse relationship between DMI and the dietary energy density observed in both the current study and those previous studies, as well as for the composition of gain observed generally in lambs fed higher energy diets (Jindal et al., 1980; Cannas et al., 2004).
The high-protein level group tended to have greater final weights and DMI than those fed low-protein diets (p = 0.10). However, the protein level did not affect the ADG or feed for gain. Some reports indicate that levels of protein play an important role on DM intake and ADG in lambs. This is particularly true when the studies included protein levels below 14%. For example, Lu and Potchoiba (1990) reported that the DM intake and ADG was influenced by dietary CP level (12, 13, and 15% CP) in a linear fashion in growing goats. Kaya et al. (2009) observed no differences in the DMI, ADG or GF when 13% and 16% CP levels were compared, yet the lambs that were fed the 10% CP diet had lower DMI, ADG and GF values than those fed 13% and 16% CP diets. In the same manner, Javed et al. (2010) reported a greater DM intake and ADG in Thalli male growing lambs that were fed a diet containing 14% of CP compared to those fed a 12% CP diet. It is thought that when the protein supply exceeds the requirement, energy becomes limiting for growth, and the animals no longer respond to additional intakes of protein (Titgemeyer, 2003). In this sense, Rocha et al. (2004) did not observe an effect of protein level (14, 16, 18, and 20% CP) on DM intake, gain and feed efficiency. Beauchemin et al. (1995) and Ruiz-Nuño et al. (2009) concluded that the protein level has a minor effect on lamb performance with regards to wool or hair sheep when finishing diets contained more than 14% of crude protein. Our findings, thus, agree with these authors.
The effects of treatments on dietary NE and observed-to-expected DMI ratios are shown in Table 3. No interactions were detected between energy and protein levels.
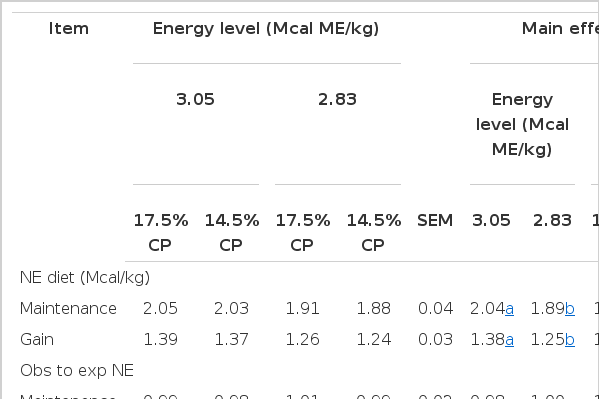
Influence of protein and energy level on dietary energy and observed to expected DMI of feedlot hair lambs
Across the entire 84 d period, the average observed-to-expected DMI of lambs was 101% of the expected value, based on tabular (NRC, 2007) estimates of dietary energy density and observed SBW and ADG values (Table 3), supporting the practicality of the prediction equations proposed by the NRC (1985) for the estimation of DMI in relation to SBW and ADG in feedlot lambs. In the same manner, the observed-to-expected NE diet averaged 0.99. This supports the theory that there was no synergistic effect resulting in a more efficient use of energy in the diet at the levels of energy and protein evaluated in this study. The extra-caloric effect of increased metabolizable protein intake due to increased protein levels in the diet could be expected in animals with high rates of growth and/or with a limited intake. Milis and Liamadis (2008) observed that CP levels above 14% in finishing diets do not affect the energy value of sheep rations, irrespective of the CP level. However, this study suggested that when the CP content is high, N degradability should be low otherwise the ME of rations is negatively affected. The percentages of rumen degradable protein (RDP) in the experimental diets used in this study were very similar among the diets and averaged 38±1.09%. Moreover, the results obtained here related to the observed-to-expected NE diet confirms that dietary effects on growth performance were mainly due to energy concentrations rather than levels of CP.
Treatment effects on the carcass characteristics and carcass composition of lambs are shown in Table 4. There was only an interaction identified between energy level and protein level for the kidney-pelvic fat (KPH) percentage (p = 0.03). With the high-energy and high-protein diet, KPH values were 22% greater than low-protein and low-energy diet. In contrast, in low energy diets there were no differences (p>0.05) in KPH proportion between protein levels.
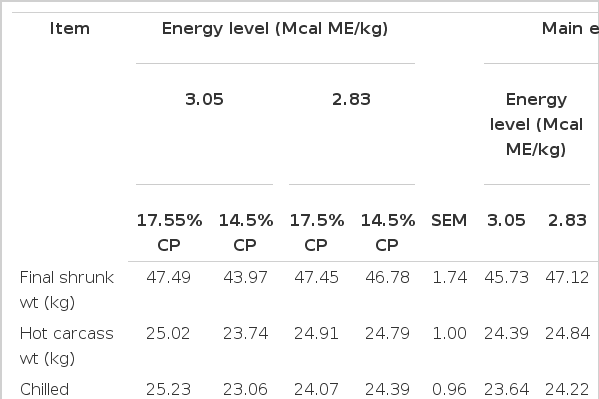
Influence of protein and energy level on carcass characteristics and carcass composition of feedlot hair lambs
In agreement with previous reports (Beuchemin et al., 1995; Rocha et al., 2004), the energy level, in high-energy finishing diets, only had small effects on HCW and dressing percentage. While, an increase of visceral fat as a result of increases of the energy density of a diet is a common response in fattening lambs (Hegarty et al., 2006; Abdel-Naset, 2009). Increasing the energy level of 2.83 to 3.05 increased the visceral fat (20.2%, p = 0.01). This increase of visceral fat is in agreement with Partida and Martinez (2010), who reported an increase of 17.1% of visceral fat when compared two energy levels (2.60 vs 2.80 Mcal/kg ME) in finishing Pelibuey lambs.
In agreement with previous reports (Beuchemin et al., 1995; Ruiz-Nuño et al., 2009), the protein level, in high-energy finishing diets, only had small effects on HCW, dressing percentage or KPH. Increasing the protein level of 14.5% to 17.5% increased the back fat thickness (22.2%, p = 0.02). Similarly, data from Ruiz-Nuño et al. (2009) show a linear increase of backfat thickness (1.42, 2.10, and 2.18 mm) in Pelibuey lambs when they were fed with 14, 16, and 18% of CP in high energy diets (2.80 Mcal/kg ME). In contrast, Ebrahimi et al. (2007) did not observe any effects of protein level (10.5, 12.5, and 14.5%) on back fat thickness in Mehraban lambs fed finishing diets contained moderated energy level (2.50 Mcal/kg ME). The increase (p<0.05) in back fat thickness promoted by the high protein level in high energy diets is a reflection of the excess of protein directed toward into fat synthesis rather than the production of lean tissue. The latter, can be supported by the absence of changes in rib eye area observed between treatments in the present study.
No interactions were detected between energy and protein levels on carcass composition. Muscle and fat averaged 58.07±0.47% and 23.17±0.24%, respectively, and were not modified by the effects of protein or energy levels. These values are in agreement with values of 55.20% and 18% reported by Partida et al. (2009) in Pelibuey×Dorset animals that were fed with a diet containing 2.80 Mcal/kg ME and harvested at an LW of 47 kg, but were lower for protein and higher for fat than those values reported of 64.3% of muscle and 13.8% of fat obtained in Pelibuey lambs harvested at an LW of 44 kg (Partida and Martínez, 2010). The differences in carcass composition observed in the present study and those reported by Partida and Martinez (2010) were mainly due to differences in the rate of growth. The average gain observed in the present study was 38% greater (280 vs 177 g/d) than those seen by Partida and Martínez (2010).
Energy levels did not affect bone percentages, but diets with high protein levels had greater bone percentages than low-protein level diets (3.3%, p<0.05). The reason for this is uncertain, since the bone formation is completed earlier than other tissues in growing animals (Colomer-Rocher et al., 1992), it is conceivable that there were no differences in bone percentages among treatments due to the likelihood of completion of bone formation before this growth trial, thus, these differences in particular may be due to measurement mistakes rather than protein level effect. Muscle:bone and muscle:fat ratios were not affected by the treatments. In contrast, Partida and Martinez (2010) observed that a higher energetic diet promoted the increase of muscle tissue and improved the muscle:fat ratio from 4.66 to 6.00 in Pelibuey lambs which were harvested at an LW of 44 kg.
It is concluded that dietary effects on growth performance and dietary energetics in hairy lambs (initial weight ~22 kg) were due mainly to energy concentrations rather than levels of CP in high energy finishing diets (i.e. 2.80 Mcal/kg ME) with a minimal concentration of 14.5% protein. Thus, when a single concentrate diet is allocated during the finishing period, there appears to be no advantage to increasing the protein concentration beyond 14.5%. In this type of diets (>2.80 Mcal/kg ME), visceral fat deposition is the main nutrient that is affected when lambs are slaughtered at similar weights, while protein and energy levels had small effects on carcass characteristics.
ACKNOWLEDGEMENTS
The authors are thankful to the SEP-PROMEP for financial support. Project: Integral Development of UASCA-205. Ref #: 2007-25-41.